1. Recall from the zoology course which parts of the brain all vertebrates have. What department are the cerebral hemispheres derived from? In which group of animals do they first appear?
All vertebrates are characterized by the presence of 5 parts of the brain: medulla hindbrain, midbrain, diencephalon, forebrain.
General overview of the structure of the cerebral hemispheres
Misunderstanding the Horse Brain It is not uncommon to hear the following statements: There are two horses in one horse: the right horse and the reliable left horse and the unpredictable right horse, thus referring to the two hemispheres of the brain: the left hemisphere for logic. You should never ride a horse using the right brain and teach the horse to use his left brain first. The horse has two visual consciousnesses. The horse does not think, he instinctively reacts. But these claims are not based on scientific evidence.
The cerebral hemispheres are derivatives of the forebrain and first appear in amphibians, but they are poorly developed, the cerebral cortex is practically absent.
2. Describe the structure hemispheres person. Draw a schematic section of the cerebral hemispheres, indicating in the figure the gray matter of the cortex, the gray matter of the nuclei, the white matter, and the ventricles.
Looking at the history of mammalian evolution into hominids and the horse's place in that history is a way to argue against these errors in the analysis of horse behavior. The mammalian class has several divisions or orders, such as rodents, carnivores, insectivores, ungulates, and primates.
Mammals are animals characterized by the production of milk from the mammary glands of the female. The horse belongs to the class of mammals and to the order of ungulates, since the edge of the fingers is covered with a hoof, and the limbs rest on the last phalanx. However, the horse was only domesticated later, the first traces of domestication dating from 500 years to a thousand years. Brain evolution in some adult mammals. total brain weight in grams.
In humans, the forebrain is represented by two hemispheres and the corpus callosum connecting the hemispheres. The large hemispheres: right and left - cover the middle and diencephalon and make up to 80% of the mass of the brain of an adult. On the surface of each hemisphere there are many furrows and convolutions. There are 4 main sulci (central, lateral and parietal-occipital) that divide each hemisphere into lobes. The surface layer consists of gray matter (cortex), below it is white matter, consisting of axons of nerve cells, whose bodies lie in the cortex or which carry information to the cells of the cortex. Large accumulations of gray matter (subcortical nuclei) and cavities (lateral ventricles) lie in the thickness of the white matter. Through the ventricles of the brain and the central canal spinal cord cerebrospinal fluid circulates, which provides nutrition to the subcortical structures.
The total weight of the brain, expressed as a percentage of body weight. Primates and hominids. Within the class of mammals there is also an order of primates that appear at the beginning of the Tertiary era. Among these primates are pongids or anthropopod apes and hominids. Hominid is a family of bipedal primates i.e. Austratopites like Lucy and later males.
Brain Evolution The study of the evolution of living things over millions of years from the first mammals to humans has made it possible to observe general trend of the brain to an increase in volume globally and in favor of specific areas of the brain. In mammals, this increase was mainly in favor of the cerebral hemispheres and cerebellum, so that, unlike birds and reptiles, only mammals developed neorctery with 6 cell layers, this nocort represents more than half the brain volume in the most advanced mammals.
3. What is the cerebral cortex? Where is it?
The surface layer of the gray matter of the cerebral hemispheres is called the cortex. The cortex consists of several layers of neuron bodies, different in structure and function. It is believed that its composition includes about 12-18 billion cells, the thickness is 1.5-4.5 mm, and the area is 1.7-2.5 thousand cm2.
The structure of the frontal lobe
Hominid evolution has also seen an increase in cranial capacity through the gradual growth of the brain and cerebellum. One of the features human brain unlike other mammals is its anatomical and functional asymmetry. As an example, large parts of the left hemisphere are specialized in the production and understanding of language, in writing and reading. The right hemisphere is more specifically associated with spatial orientation, in detecting visual forms and musical knowledge. The left hemisphere is more dominant for processes, logical, analytical and right for holistic processes.
4. Explain the significance of the furrows and convolutions on the surface of the cerebral hemispheres.
Furrows and gyrus significantly increase the surface of the cerebral cortex, according to some sources up to 10-12 times.
5. What is the function of the white matter of the cerebral hemispheres?
The white matter forms pathways connecting the cortex and the cortex with the rest of the nervous system.
Thus, brain asymmetry is feature human, as well as the lack of a remarkable network, in monkeys, still having symmetrical brains, the ancient norcort retaining its symmetrical functioning, so logic should not be implied. in the horse one of the two hemispheres is in a certain function, some of the new masters say.
But even in humans, the cerebral hemisphere is never the only one that interferes with the function, but it only seems to be more connected, implicit in this function. In fact, the basic functions of the brain remain largely distributed across all two hemispheres of the brain. In this regard, it is also important to recall the presence of a corpus callosum structure in both humans and horses. This structure is real connection between the two hemispheres that communicate with each other. To say, therefore, that the connection between the hemispheres of the brain on a horse is absolutely incorrect.
6. What lobes are isolated in the cerebral hemispheres?
In the hemispheres, the frontal, 2 parietal, 2 temporal and occipital lobes are distinguished
7. Distinguish between the concepts of "lobes of the cerebral hemispheres" and "zones of the cerebral hemispheres." Give examples when they match, do not match.
The lobes of the cerebral hemispheres are the division of the surface of the cortex according to the anatomical principle: in each hemisphere, the frontal, occipital, parietal, and temporal lobes are distinguished.
Cortical zones - a section of the cerebral cortex, characterized by the uniformity of the structure and functions performed.
In our brain, lobes and zones do not coincide; one lobe consists of several zones. The occipital lobe consists of areas of vision and visual recognition. In the temporal lobe and next to it are the zones of smell, hearing and taste.
8. When examining a blind patient, it was found that his eyes and optic nerves were not damaged. Why can't he see?
The inability to see can be caused, in addition to damage to the eyes or optic nerves, and damage to the visual zones of the occipital lobes of the cerebral hemispheres.
9. Using additional sources of information, find out if the functions of the left and right hemispheres of the brain differ.
There is a "functional asymmetry" between the right and left hemispheres of the brain, that is, their functions are different. This was proved during experiments on the cutting of communication lines between the hemispheres (subsequently, such operations began to be carried out for medical reasons in certain diseases, such as Parkinson's disease). Right-handed people have the left hemisphere, while left-handed people have the right hemisphere. The right hemisphere is responsible for creative thinking, forms the basis of creativity, making non-standard decisions; The visual area of the right hemisphere is responsible for face recognition. The left hemisphere provides logical reasoning and abstract thinking, it contains the centers of oral and written speech, the formation of decisions; the visual zone is responsible for recognizing letters and numbers. Therefore, jokingly, right-handers are called mathematicians, and left-handers are artists.
10. There are everyday concepts of "male logic" and "female logic". Are there any grounds for such differences?
In the external and internal structure, there is no difference between the brains of men and women, but, despite the fact that the structure of our brains is almost the same, we are individual. The main differences in the perception of the world according to the “male” or “female” type are laid down in early childhood depending on our upbringing and under the influence of hormones of the gonads during growing up, but all of them are only psychological in nature.
14.1. GENERAL PROVISIONS
End brain (telencephalon), or big brain (cerebrum), located in the supratentorial space of the cranial cavity consists of two large
hemispheres (gemispherium cerebralis),separated by a deep longitudinal slit (fissura longitudinalis cerebri), in which the crescent of the brain is immersed (falx cerebri) representing a duplication of the dura mater. The large hemispheres of the brain make up 78% of its mass. Each of the cerebral hemispheres has lobes: frontal, parietal, temporal, occipital and limbic. They cover the structures of the diencephalon and the brain stem and cerebellum located below the cerebellar mantle (subtentorially).
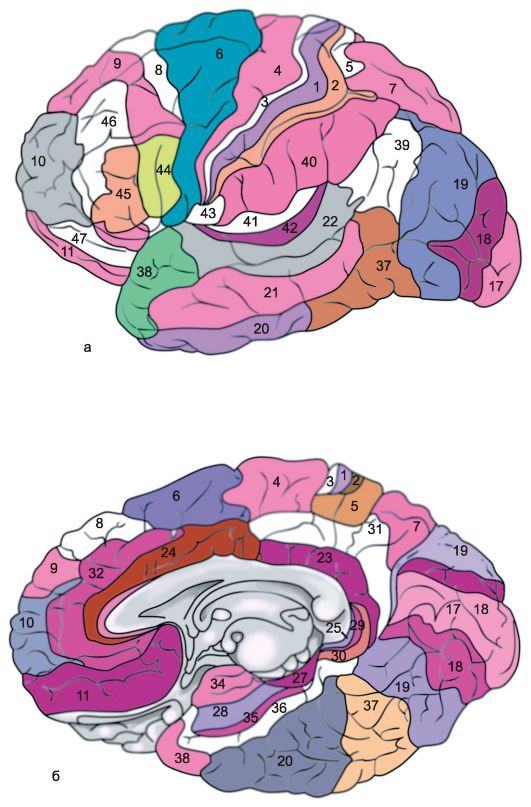
Each of the cerebral hemispheres has three surfaces: upper lateral, or convexital (Fig. 14.1a), - convex, facing the bones of the cranial vault; internal (Fig. 14.1b), adjacent to the large falciform process, and lower, or basal (Fig. 14.1c), repeating the relief of the base of the skull (anterior and middle pits) and the cerebellar tenon. In each hemisphere, three edges are distinguished: upper, lower inner and lower outer, and three poles: anterior (frontal), posterior (occipital) and lateral (temporal).
The cavity of each cerebral hemisphere is lateral ventricle of the brain while the left lateral ventricle is recognized as the first, the right - the second. The lateral ventricle has a central part located deep in the parietal lobe (lobus parietalis) and three horns extending from it: the anterior horn penetrates the frontal lobe (lobus frontalis), lower - to the temporal (lobus temporalis), posterior - in the occipital (lobus occipitalis). Each of the lateral ventricles communicates with the third ventricle of the brain through the interventricular hole Monroe.
The central sections of the medial surface of both hemispheres are interconnected by cerebral commissures, the most massive of which is the corpus callosum, and structures of the diencephalon.
The telencephalon, like other parts of the brain, consists of gray and white matter. Gray matter is located in the depths of each hemisphere, forming subcortical nodes there, and along the periphery of the free surfaces of the hemisphere, where it makes up the cerebral cortex.
The main issues related to the structure, functions of the basal ganglia and variants of the clinical picture when they are affected are discussed in chapters 5, 6. The cerebral cortex is approximately
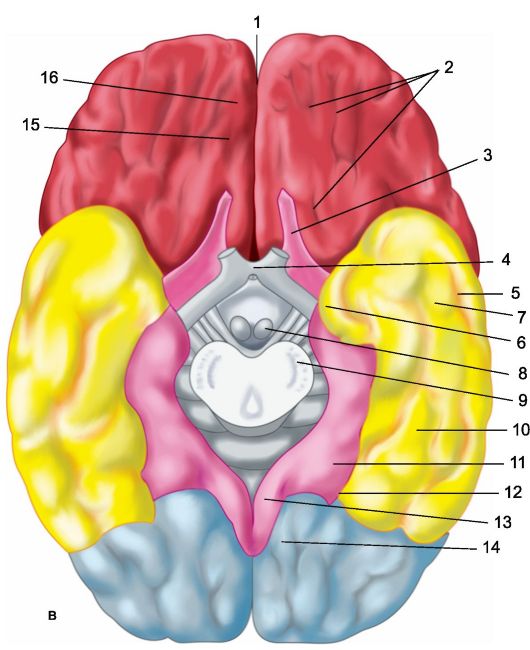
Rice. 14.1.Hemispheres of the brain.
a - upper lateral surface of the left hemisphere: 1 - central sulcus; 2 - orbital part of the lower frontal gyrus; I - frontal lobe; 3 - precentral gyrus; 4 - precentral furrow; 5 - superior frontal gyrus; 6 - middle frontal gyrus; 7 - tegmental part of the inferior frontal gyrus; 8 - lower frontal gyrus; 9 - lateral furrow; II - parietal lobe: 10 - postcentral gyrus; 11 - postcentral furrow; 12 - intraparietal groove; 13 - supramarginal gyrus; 14 - angular gyrus; III - temporal lobe: 15 - superior temporal gyrus; 16 - upper temporal sulcus; 17 - middle temporal gyrus; 18 - middle temporal sulcus; 19 - lower temporal gyrus; IV - occipital lobe: b - medial surface of the right hemisphere: 1 - paracentral lobule, 2 - precuneus; 3 - parieto-occipital sulcus; 4 - wedge, 5 - lingual gyrus; 6 - lateral occipitotemporal gyrus; 7 - parahippocampal gyrus; 8 - hook; 9 - vault; 10 - corpus callosum; 11 - superior frontal gyrus; 12 - cingulate gyrus; c - lower surface of the cerebral hemispheres: 1 - longitudinal interhemispheric fissure; 2 - orbital furrows; 3 - olfactory nerve; 4 - optic chiasm; 5 - middle temporal sulcus; 6 - hook; 7 - lower temporal gyrus; 8 - mastoid body; 9 - base of the brain stem; 10 - lateral occipitotemporal gyrus; 11 - parahippocampal gyrus; 12 - collateral groove; 13 - cingulate gyrus; 14 - lingual gyrus; 15 - olfactory groove; 16 - direct gyrus.
3 times the surface of the hemispheres visible during external examination. This is due to the fact that the surface of the cerebral hemispheres is folded, has numerous depressions - furrows (sulci cerebri) and located between them convolutions (gyri cerebri). The cerebral cortex covers the entire surface of the convolutions and furrows (hence its other name is pallium - a cloak), while sometimes penetrating to a great depth into the substance of the brain.
The severity and location of the furrows and convolutions of the cerebral hemispheres are variable to a certain extent, however, the main ones are formed in the process of ontogenesis and are constant, characteristic of each normally developed brain.
14.2. MAJOR GROOCHES AND GRIPS OF THE HEMISPHERES OF THE BRAIN
Upper lateral (convexital) surface of the hemispheres (Fig. 14.1a). The largest and deepest lateral furrow (sulcus lateralis),or sylvian furrow, - separates the frontal and anterior parts of the parietal lobe from the temporal lobe located below. The frontal and parietal lobes are separated central, or Roland, furrow(sulcus centralis), which cuts through the upper edge of the hemisphere and goes down and forward along its convexital surface, slightly short of the lateral groove. The parietal lobe is separated from the occipital lobe located behind it by the parietal-occipital and transverse occipital grooves passing along the medial surface of the hemisphere.
In the frontal lobe in front of the central gyrus and parallel to it is the precentral (gyrus precentralis), or anterior central, gyrus, which is bounded anteriorly by the precentral sulcus (sulcus precentralis). The superior and inferior frontal grooves depart anteriorly from the precentral sulcus, dividing the convexital surface of the anterior sections of the frontal lobe into three frontal gyrus - superior, middle and inferior (gyri frontales superior, media et inferior).
The anterior section of the convexital surface of the parietal lobe is located behind the central sulcus postcentral (gyrus postcentralis), or posterior central, gyrus. Behind it is bordered by the postcentral sulcus, from which the intraparietal sulcus stretches back. (sulcus intraparietalis), separating the superior and inferior parietal lobules (lobuli parietales superior et inferior). In the lower parietal lobule, in turn, the supramarginal gyrus is distinguished (gyrus supramarginalis), surrounding the posterior part of the lateral (Sylvian) groove, and the angular gyrus (girus angularis), bordering the back of the superior temporal gyrus.
On the convexital surface of the occipital lobe of the brain, the furrows are shallow and can vary significantly, as a result of which the nature of the convolutions located between them is also variable.
The convexital surface of the temporal lobe is divided by the superior and inferior temporal sulci, which are almost parallel to the lateral (Sylvian) sulcus, dividing the convexital surface of the temporal lobe into the superior, middle, and inferior temporal gyri (gyri temporales superior, media et inferior). The superior temporal gyrus forms the inferior lip of the lateral (Sylvian) sulcus. On its surface facing
side of the lateral furrow, there are several transverse small furrows, highlighting small transverse gyrus on it (gyrus of Geschl), which can be seen only by spreading the edges of the lateral furrow.
The anterior part of the lateral (Sylvian) groove is a depression with a wide bottom, forming the so-called island (insula) or insular lobe (lubus insularis). The upper edge of the lateral furrow covering this island is called tire (operculum).
Inner (medial) surface of the hemisphere (Fig. 14.1b). The central part of the inner surface of the hemisphere is closely connected with the structures of the diencephalon, from which it is delimited by those related to the large brain vault (fornix) and corpus callosum (corpus callosum). The latter is bordered on the outside by a furrow of the corpus callosum (sulcus corporis callosi), starting at the front of it - the beak (rostrum) and ending at its thickened rear end (splenium). Here, the sulcus of the corpus callosum passes into the deep hippocampal sulcus (sulcus hippocampi), which penetrates deep into the substance of the hemisphere, pressing it into the cavity of the lower horn of the lateral ventricle, as a result of which the so-called ammonium horn is formed.
Somewhat departing from the sulcus of the corpus callosum and the hippocampal sulcus, the corpus callosum, subparietal and nasal sulci are located, which are a continuation of each other. These grooves delimit from the outside the arcuate part of the medial surface of the cerebral hemisphere, known as limbic lobe(lobus limbicus). There are two convolutions in the limbic lobe. The upper part of the limbic lobe is the superior limbic (superior marginal), or girdle, gyrus (girus cinguli), the lower part is formed by the inferior limbic gyrus, or seahorse gyrus (girus hippocampi), or parahippocampal gyrus (girus parahypocampalis), in front of which there is a hook (uncus).
Around the limbic lobe of the brain are the formations of the inner surface of the frontal, parietal, occipital and temporal lobes. Most of the inner surface of the frontal lobe is occupied by the medial side of the superior frontal gyrus. On the border between the frontal and parietal lobes of the cerebral hemisphere is located paracentral lobule (lobulis paracentralis), which is, as it were, a continuation of the anterior and posterior central gyri on the medial surface of the hemisphere. On the border between the parietal and occipital lobes, the parietal-occipital sulcus is clearly visible. (sulcus parietooccipitalis). From the bottom of it departs back spur furrow (sulcus calcarinus). Between these deep furrows is a triangular gyrus, known as a wedge. (cuneus). In front of the wedge is a quadrangular gyrus, related to the parietal lobe of the brain, the precuneus.
Inferior surface of the hemisphere (Fig. 14.1c). The lower surface of the cerebral hemisphere consists of formations of the frontal, temporal and occipital lobes. The part of the frontal lobe adjacent to the midline is the direct gyrus (girus rectus). Outside, it is delimited by the olfactory groove (sulcus olfactorius), to which the formations of the olfactory analyzer are adjacent from below: the olfactory bulb and the olfactory tract. Lateral to it, up to the lateral (Sylvian) groove, which extends to the lower surface of the frontal lobe, there are small orbital gyri (gyri orbitalis). The lateral sections of the lower surface of the hemisphere behind the lateral sulcus are occupied by the inferior temporal gyrus. Medial to it is the lateral temporo-occipital gyrus. (gyrus occipitotemporalis lateralis), or fusiform groove. Before-
its inner parts border on the gyrus of the hippocampus, and the posterior ones - on the lingual (gyrus lingualis) or medial temporoccipital gyrus (gyrus occipitotemporalis medialis). The latter, with its posterior end, is adjacent to the spur groove. The anterior sections of the fusiform and lingual gyri belong to the temporal lobe, and the posterior sections to the occipital lobe of the brain.
14.3. WHITE MATTER OF THE GREAT HEMISPHERES
The white matter of the cerebral hemispheres consists of nerve fibers, mainly myelin, that make up pathways that provide connections between the neurons of the cortex and clusters of neurons that form the thalamus, subcortical nodes, and nuclei. The main part of the white matter of the cerebral hemispheres is located in its depth semi-oval center, or radiant crown (corona radiata), consisting mainly of afferent and efferent projection pathways connecting the cerebral cortex with subcortical nodes, nuclei and reticular substance of the diencephalon and brain stem, with segments of the spinal cord. They are especially compactly located between the thalamus and the subcortical nodes, where they form the internal capsule described in Chapter 3.
Nerve fibers that connect parts of the cortex of one hemisphere are called associative. The shorter these fibers and the connections they form, the more superficial they are; longer associative connections, located deeper, connect relatively distant parts of the cerebral cortex (Fig. 14.2 and 14.3).
The fibers that connect the cerebral hemispheres and therefore have a common transverse orientation are called commissural, or sleeping. Commissural fibers connect identical parts of the cerebral hemispheres, creating the possibility of combining their functions. They form three spikes large brain: the most massive of them - corpus callosum (corpus callosum), in addition, commissural fibers make up anterior commissure, located under the beak of the corpus callosum (rostrum corporis collosum) and connecting both olfactory regions, as well as commissure of the vault (commissura fornicis), or a hippocampal commissure formed by fibers connecting the structures of the ammon horns of both hemispheres.
In the anterior part of the corpus callosum there are fibers connecting the frontal lobes, then there are fibers connecting the parietal and temporal lobes, the posterior part of the corpus callosum connects the occipital lobes of the brain. The anterior commissure and commissure of the fornix mainly unite sections of the ancient and old cortex of both hemispheres; the anterior commissure, in addition, provides a connection between their middle and lower temporal gyri.
14.4. Olfactory system
In the process of phylogenesis, the development of the large brain is associated with the formation of the olfactory system, the functions of which contribute to the preservation of the viability of animals and are of no small importance for human life.
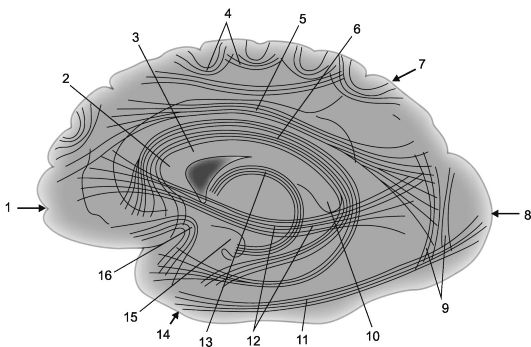
Rice. 14.2.Associative cortical-cortical connections in the cerebral hemispheres [according to V.P. Vorobyov].
1 - frontal lobe; 2 - knee of the corpus callosum; 3 - corpus callosum; 4 - arcuate fibers; 5 - upper longitudinal beam; 6 - cingulate gyrus; 7 - parietal lobe, 8 - occipital lobe; 9 - vertical bundles of Wernicke; 10 - roller of the corpus callosum;
11 - lower longitudinal beam; 12 - subcausal bundle (frontal-occipital lower bundle); 13 - vault; 14 - temporal lobe; 15 - hook of the gyrus of the hippocampus; 16 - hook bundles (fasciculus uncinatus).
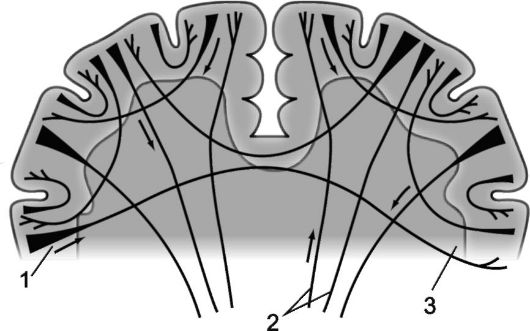
Rice. 14.3.Myeloarchitectonics of the cerebral hemispheres.
1 - projection fibers; 2 - commissural fibers; 3 - associative fibers.
14.4.1. The structure of the olfactory system
The bodies of the first neurons of the olfactory system are located in the mucous membrane nose, mainly upper part of the nasal septum and upper nasal passage. Olfactory cells are bipolar. Their dendrites come to the surface of the mucous membrane and end here with specific receptors, and axons are grouped in the so-called olfactory filaments (filiolfactorii), the number of which on each side is about twenty. Such a bundle of olfactory filaments and makes up the I cranial, or olfactory, nerve(Fig. 14.4). These threads pass into the anterior (olfactory, olfactory) cranial fossa through the ethmoid bone and end at cells located here olfactory bulbs. The olfactory bulbs and the proximal olfactory tracts are, in fact, a consequence of the protrusions of the substance of the large brain formed in the process of ontogenesis and represent structures related to it.
The olfactory bulbs contain cells that are the bodies of the second neurons. olfactory pathway, whose axons form olfactory tracts (tracti olfactorii), located under the olfactory grooves, lateral to the direct convolutions located on the basal surface of the frontal lobes. Olfactory tracts are directed backward to the subcortical olfactory centers. Approaching the anterior perforated plate, the fibers of the olfactory tract are divided into medial and lateral bundles, forming an olfactory triangle on each side. Later, these fibers are suitable to the bodies of the third neurons of the olfactory analyzer, located

Rice. 14.4.Olfactory analyzer.
1 - olfactory cells; 2 - olfactory threads (in total they make up the olfactory nerves); 3 - olfactory bulbs; 4 - olfactory tracts; 5 - olfactory triangles; 6 - parahippocampal gyrus; 7 - projection zone of the olfactory analyzer (simplified diagram).
in the perialmond-shaped and subcallosal areas, in the nuclei of the transparent septum, located anterior to the anterior commissure. The anterior commissure connects both olfactory regions and also provides their connection to the limbic system of the brain. Part of the axons of the third neurons of the olfactory analyzer, passing through the anterior commissure of the brain, crosses.
Axons of third neurons olfactory analyzer, located in the subcortical olfactory centers, heading towards phylogenetically old crust mediobasal surface of the temporal lobe (to the piriform and parahippocampal gyrus and to the hook), where the projection olfactory zone is located, or the cortical end of the olfactory analyzer (field 28, according to Brodmann).
The olfactory system is thus the only sensory system in which specific impulses bypass the thalamus on their way from the receptors to the cortex. However, The olfactory system has especially pronounced connections with the limbic structures of the brain, and the information received through it has a significant impact on the state of the emotional sphere and the functions of the autonomic nervous system. Smells can be pleasant and unpleasant, they affect appetite, mood, can cause a variety of vegetative reactions, in particular nausea, vomiting.
14.4.2. Investigation of the sense of smell and the significance of its disorders for topical diagnostics
When examining the state of smell, it is necessary to find out whether the patient smells, whether these sensations are the same on both sides, whether the patient differentiates the nature of the smells felt, whether he has olfactory hallucinations - paroxysmal sensations of smell that are absent in the environment.
To study the sense of smell, odorous substances are used, the smell of which is not sharp (pungent odors can cause irritation of the trigeminal nerve receptors located in the nasal mucosa) and is known to the patient (otherwise it is difficult to recognize the perversion of smell). The sense of smell is checked on each side separately, while the other nostril must be closed. You can use specially prepared sets of weak solutions of odorous substances (mint, tar, camphor, etc.), in practical work, improvised means (rye bread, soap, banana, etc.) can also be used.
Decreased sense of smell - hyposmia, lack of smell - anosmia, heightened sense of smell - hyperosmia, perversion of odors dysosmia, sensation of smell in the absence of a stimulus - parosmia, subjective sensation of an unpleasant odor that actually exists and is caused by organic pathology in the nasopharynx - kakosmiya, odors that do not really exist, which the patient feels paroxysmally - olfactory hallucinations - are more often the olfactory aura of temporal lobe epilepsy, which may be due to different reasons, in particular a tumor of the temporal lobe.
Hyposmia or anosmia on both sides is usually the result of damage to the nasal mucosa due to acute catarrh, influenza, allergic rhinitis, atrophy of the mucous membrane
nose due to chronic rhinitis and prolonged use of vasoconstrictor nasal drops. Chronic rhinitis with atrophy of the nasal mucosa (atrophic rhinitis), Sjögren's disease dooms a person to persistent anosmia. Bilateral hyposmia can be caused by hypothyroidism, diabetes mellitus, hypogonadism, renal failure, prolonged contact with heavy metals, formaldehyde, etc.
However, unilateral hyposmia or anosmia is often the result of an intracranial tumor, more often meningioma of the anterior cranial (olfactory) fossa, which accounts for up to 10% of intracranial meningiomas, as well as some glial tumors of the frontal lobe. Olfactory disorders occur as a result of compression of the olfactory tract on the side of the pathological focus and may be the only focal symptom of the disease for a certain time. Tumors can be visualized by CT or MRI scanning. As the meningioma of the olfactory fossa increases, as a rule, mental disorders characteristic of the frontal syndrome develop (see Chapter 15).
Unilateral damage to the parts of the olfactory analyzer located above its subcortical centers, due to incomplete decussation of the pathways at the level of the anterior cerebral commissure, usually does not lead to a significant decrease in the sense of smell. Irritation by the pathological process of the cortex of the mediobasal parts of the temporal lobe, primarily the parahippocampal gyrus and its hook, can cause a paroxysmal occurrence olfactory hallucinations. The patient suddenly begins to smell for no reason, often of an unpleasant nature (the smell of burnt, rotten, rotten, burnt, etc.). Olfactory hallucinations in the presence of an epileptogenic focus in the mediobasal regions of the temporal lobe of the brain may be a manifestation of the aura of an epileptic seizure. The defeat of the proximal part, in particular the cortical end of the olfactory analyzer, can cause moderate bilateral (more on the opposite side) hyposmia and impaired ability to identify and differentiate odors (olfactory agnosia). The last form of olfactory disorder, which manifests itself in old age, is most likely associated with a violation of the function of the cortex due to atrophic processes in its projection olfactory zone.
14.5. LIMBIC-RETICULAR COMPLEX
In 1878 P. Broca(Broca P., 1824-1880) under the name "large marginal, or limbic, lobe" (from lat. limbus - edge) united the hippocampus and the cingulate gyrus, interconnected by means of the isthmus of the cingulate gyrus, located above the ridge of the corpus callosum.
In 1937 D. Papets(Papez J.), on the basis of experimental data, put forward a reasoned objection to the previously existing concept of the involvement of the mediobasal structures of the cerebral hemispheres mainly in the provision of smell. He suggested that the main part of the mediobasal parts of the cerebral hemisphere, then called the olfactory brain (rhinencephalon), to which the limbic lobe belongs, is the morphological basis of the nervous mechanism of affective behavior, and united them under the name"emotional circle" which included the hypothalamus,
anterior nuclei of the thalamus, cingulate gyrus, hippocampus and their connections. Since then, these structures have also been referred to by physiologists as around Papez.
concept "visceral brain" suggested P.D. McLean (1949), thus denoting a complex anatomical and physiological association, which since 1952 has been called "limbic system". Later it turned out that the limbic system is involved in the performance of diverse functions, and now most of it, including the cingulate and hippocampal (parahippocampal) gyrus, is usually combined into the limbic region, which has numerous connections with the structures of the reticular formation, making up with it limbic-reticular complex wide circle physiological and psychological processes.
Currently to limbic lobe it is customary to attribute elements of the old cortex (archiocortex), covering the dentate gyrus and the hippocampal gyrus; ancient cortex (paleocortex) of the anterior hippocampus; as well as the middle, or intermediate, cortex (mesocortex) of the cingulate gyrus. Term "limbic system" includes components of the limbic lobe and related structures - entorhinal (occupying most of the parahippocampal gyrus) and septal regions, as well as the amygdala complex and mastoid body (Duus P., 1995).
Mastoid body connects the structures of this system with the midbrain and with the reticular formation. Impulses originating in the limbic system can be transmitted through the anterior nucleus of the thalamus to the cingulate gyrus and to the neocortex along pathways formed by associative fibers. Impulses originating in the hypothalamus can reach the orbitofrontal cortex and the medial dorsal nucleus of the thalamus.
Numerous direct and feedback provide interconnection and interdependence of limbic structures and many formations of the diencephalon and oral parts of the brainstem (nonspecific nuclei of the thalamus, hypothalamus, putamen, frenulum, reticular formation of the brainstem), as well as with subcortical nuclei (pallidus, putamen, caudate nucleus) and with the new cortex cerebral hemispheres, primarily with the cortex of the temporal and frontal lobes.
Despite phylogenetic, morphological, and cytoarchitectonic differences, many of the mentioned structures (limbic region, central and medial structures of the thalamus, hypothalamus, brainstem reticular formation) are usually included in the so-called limbic-reticular complex, which acts as a zone of integration of many functions, providing the organization of polymodal, holistic reactions of the body to various influences, which is especially pronounced in stressful situations.
The structures of the limbic-reticular complex have a large number of entrances and exits through which vicious circles of numerous afferent and efferent connections pass, ensuring the combined functioning of the formations included in this complex and their interaction with all parts of the brain, including the cerebral cortex.
In the structures of the limbic-reticular complex, there is a convergence of sensitive impulses that occur in intero- and exteroreceptors, including the receptor fields of the sense organs. On this basis, in the limbic-reticular complex, primary synthesis of information about the state of the internal environment of the body, as well as about the factors affecting the body external environment, and elementary needs, biological motivations and accompanying emotions are formed.
The limbic-reticular complex determines the state of the emotional sphere, participates in the regulation of vegetative-visceral relationships aimed at maintaining the relative constancy of the internal environment (homeostasis), as well as energy supply and correlation of motor acts. The level of consciousness, the possibility of automated movements, the activity of motor and mental functions, speech, attention, the ability to orientate, memory, the change of wakefulness and sleep depend on its state.
Damage to the structures of the limbic-reticular complex can be accompanied by a variety of clinical symptoms: pronounced changes in the emotional sphere of a permanent and paroxysmal nature, anorexia or bulimia, sexual disorders, memory impairment, in particular signs of Korsakoff's syndrome, in which the patient loses the ability to remember current events (retains current events in memory for no more than 2 minutes), autonomic-endocrine disorders, sleep disorders, psychosensory disorders in the form of illusions and hallucinations, changes in consciousness, clinical manifestations of akinetic mutism, epileptic seizures.
To date, a large number of studies have been conducted on the study of morphology, anatomical relationships, the function of the limbic region and other structures included in the limbic-reticular complex, however, the physiology and features of the clinical picture of its lesion today still largely need to be clarified. Most of the information about its function, especially the functions of the parahippocampal region, obtained in animal experiments methods of irritation, extirpation or stereotaxis. Obtained in this way results require caution when extrapolating to humans. Special meaning have clinical observations of patients with lesions of the mediobasal regions of the cerebral hemisphere.
In the 50-60s of the XX century. during the development of psychosurgery, there were reports of the treatment of patients with incurable mental disorders and chronic pain syndrome by bilateral cingulotomy (dissection of the cingulate gyrus), while regression of anxiety, obsessional states, psychomotor agitation, pain syndromes was usually noted, which was recognized as evidence of the involvement of the cingulate gyrus in the formation of emotions and pain. At the same time, bicingulotomy led to profound personality disorders, to disorientation, a decrease in the criticality of one's condition, and euphoria.
An analysis of 80 verified clinical cases of hippocampal lesions on the basis of the Neurosurgical Institute of the Russian Academy of Medical Sciences is given in the monograph by N.N. Bragina (1974). The author comes to the conclusion that temporal mediobasal syndrome includes viscerovegetative, motor and mental disorders, usually manifested in a complex. All the variety of clinical manifestations of N.N. Bragin reduces to two main multifactorial variants of pathology with a predominance of "irritative" and "inhibitory" phenomena.
The first of these includes emotional disorders accompanied by motor anxiety (increased excitability, verbosity, fussiness, a feeling of internal anxiety), paroxysms of fear, vital anguish, various viscerovegetative disorders (changes in pulse, respiration, gastrointestinal disorders, fever, increased sweating and etc.). In these patients, against the background of constant motor restlessness, attacks of motor excitation often occurred.
niya. The EEG of this group of patients was characterized by mild cerebral changes towards integration (accelerated and pointed alpha rhythm, diffuse beta oscillations). Repeated afferent stimuli elicited clear EEG responses, which, unlike normal ones, did not fade as stimuli were repeatedly presented.
The second (“inhibitory”) variant of the mediobasal syndrome is characterized by emotional disturbances in the form of depression with motor retardation (suppressed mood, impoverishment and slowing of the pace of mental processes, changes in motor skills, resembling the akinetic-rigid syndrome type. Viscerovegetative paroxysms noted in the first group are less characteristic. The EEG of patients in this group was characterized by cerebral changes, manifested in the predominance of slow forms of activity (irregular, slow alpha rhythm, groups of theta oscillations, diffuse delta waves).A sharp decrease in EEG reactivity attracted attention.
Between these two extreme variants there were also intermediate ones with transitional and mixed combinations of individual symptoms. So, some of them are characterized by relatively weak signs of agitated depression with increased motor activity and fatigue, with a predominance of senestopathic sensations, suspicion, which in some patients reaches paranoid states, and hypochondriacal delirium. The other intermediate group was distinguished by the extreme intensity of depressive symptoms against the background of the patient's stiffness.
These data allow us to speak about the dual (activating and inhibitory) influence of the hippocampus and other structures of the limbic region on behavioral reactions, emotions, mental status, and bioelectrical activity of the cortex. Currently, complex clinical syndromes of this type should not be regarded as primary focal. Rather, they should be considered in the light of ideas about multilevel system organization of brain activity.
S.B. Buklina (1997) cited data from a survey of 41 patients with arteriovenous malformations in the area of the cingulate gyrus. Before surgery, 38 patients had memory disorders in the forefront, and five of them had signs of Korsakoff's syndrome, in three patients Korsakoff's syndrome arose after surgery, while the severity of the increase in memory defects correlated with the degree of destruction of the cingulate gyrus itself, as well as with involvement in pathological process of the adjacent structures of the corpus callosum, while the amnesic syndrome did not depend on the side of the malformation location and its localization along the length of the cingulate gyrus.
The main characteristics of the identified amnestic syndromes were disorders in the reproduction of auditory-speech stimuli, violations of the selectivity of traces in the form of inclusions and contaminations, and inability to retain meaning in the transmission of a story. In most patients, the criticality of assessing their condition was reduced. The author noted the similarity of these disorders with amnestic defects in patients with frontal lesions, which can be explained by the presence of connections between the cingulate gyrus and the frontal lobe.
More widespread pathological processes in the limbic region cause pronounced disorders of the vegetative-visceral functions.
corpus callosum(corpus callosum)- the largest commissure between the cerebral hemispheres. Its anterior sections, in particular the knee of the corpus callosum
body (genu corporis callosi), connect the frontal lobes, the middle sections - the trunk of the corpus callosum (truncus corporis callosi)- provide communication between the temporal and parietal sections of the hemispheres, the posterior sections, in particular the corpus callosum ridge (splenium corporis callosi), connect the occipital lobes.
Lesions of the corpus callosum are usually accompanied by disorders of the mental state of the patient. The destruction of its anterior section leads to the development of the “frontal psyche” (aspontaneity, violations of the action plan, behavior, criticism, characteristic of frontal callous syndrome - akinesia, amimia, aspontaneity, astasia-abasia, apraxia, grasping reflexes, dementia). Disconnection of connections between the parietal lobes leads to perversion understanding "body plans" and appearance of apraxia mostly in the left hand. Dissociation of the temporal lobes may manifest violation of the perception of the external environment, the loss of the correct orientation in it (amnestic disorders, confabulations, the syndrome of what has already been seen etc.). Pathological foci in the posterior parts of the corpus callosum are often characterized by signs of visual agnosia.
14.6. ARCHITECTONICS OF THE BRAIN CORTEX
The structure of the cerebral cortex is heterogeneous. Less complex in structure, early emerging in the process of phylogenesis ancient bark (archiocortex) and old bark (paleocortex), related basically to limbic lobe brain. The greater part of the cerebral cortex (95.6%), due to its later phylogenetic formation, is called new bark (neocortex) and has a much more complex multilayer structure, but also heterogeneous in its various zones.
Due to the fact that the architectonics of the cortex is in a certain connection with its function, much attention has been devoted to its study. One of the founders of the doctrine of the cytoarchitectonics of the cortex was V.A. Betz (1834-1894), who for the first time in 1874 described the large pyramidal cells of the motor cortex (Betz cells) and determined the principles for dividing the cerebral cortex into main areas. In the future, a great contribution to the development of the theory of the structure of the cortex was made by many researchers - A. Campbell (A. Cambell), E. Smith (E. Smith), K. Brodmann (K. Brodmann), Oscar Vogt and Cecilia Vogt (O. Vogt , S. Vogt). Great merit in the study of the architectonics of the cortex belongs to the staff of the Institute of the Brain of the Academy of Medical Sciences (S.A. Sarkisov, N.I. Filimonov, E.P. Kononova, etc.).
The main type of structure of the new crust (Fig. 14.5), with which all its sections are compared - a cortex consisting of 6 layers (homotypic cortex, according to Brodman).
Layer I - molecular, or zonal, the most superficial, poor in cells, its fibers have a direction, mainly parallel to the surface of the cortex.
II layer - outer granular. Consists of a large number of densely arranged small granular nerve cells.
III layer - small and medium pyramids, the widest. It consists of pyramidal cells, the sizes of which are not the same, which allows dividing this layer into sublayers in most cortical fields.
IV layer - internal granular. It consists of densely arranged small cells-grains of a round and angular shape. This layer is the most variable
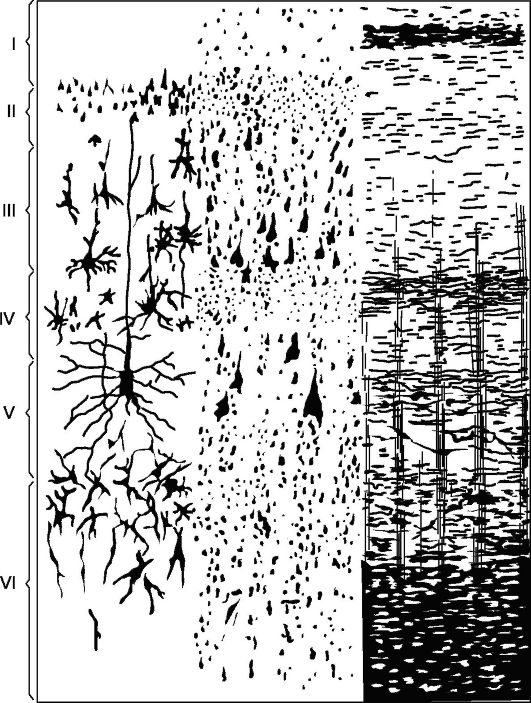
Rice. 14.5.Cytoarchitectonics and myeloarchitectonics of the motor zone of the cerebral cortex.
Left: I - molecular layer; II - outer granular layer; III - layer of small and medium pyramids; IV - inner granular layer; V - layer of large pyramids; VI - layer of polymorphic cells; on the right - elements of myeloarchitectonics.
in some fields (for example, field 17), it is divided into sublayers, in some places it sharply becomes thinner and even completely disappears.
V layer - large pyramids, or ganglionic. Contains large pyramidal cells. In some areas of the brain, the layer is divided into sublayers, in the motor zone it consists of three sublayers, the middle of which contains Betz's giant pyramidal cells, reaching 120 microns in diameter.
VI layer - polymorphic cells, or multiform. Consists mainly of triangular spindle-shaped cells.
The structure of the cerebral cortex has a large number of variations due to changes in the thickness of individual layers, thinning or disappearance or,
on the contrary, thickening and division into sublayers of some of them (heterotypic zones, according to Brodman).
The cortex of each cerebral hemisphere is divided into several regions: occipital, superior and inferior parietal, postcentral, central gyri, precentral, frontal, temporal, limbic, insular. Each of them in accordance with the characteristics subdivided into a number of fields, moreover, each field has its own conventional ordinal designation (Fig. 14.6).
The study of the architectonics of the cerebral cortex, along with physiological, including electrophysiological, studies and clinical observations, greatly contributed to the solution of the problem of the distribution of functions in the cortex.
14.7. PROJECTION AND ASSOCIATION FIELDS OF THE CORTUS
In the process of developing the doctrine of the role of the cerebral cortex and its individual sections in the performance of certain functions, there were different, sometimes opposite, points of view. Thus, there was an opinion about a strictly local representation in the cerebral cortex of all human abilities and functions, up to the most complex, mental (localizationism, psychomorphologism). He was opposed by another opinion about the absolute functional equivalence of all parts of the cerebral cortex (equipotentialism).
An important contribution to the theory of localization of functions in the cerebral cortex was made by I.P. Pavlov (1848-1936). He singled out the projection zones of the cortex (the cortical ends of the analyzers certain types sensitivity) and the associative zones located between them, studied the processes of inhibition and excitation in the brain, their influence on the functional state of the cerebral cortex. The division of the cortex into projection and associative zones contributes to understanding the organization of the work of the cerebral cortex and justifies itself in solving practical problems, in particular, in topical diagnostics.
projection zones provide mainly simple specific physiological acts, primarily the perception of sensations of a certain modality. The projection pathways approaching them connect these zones with the receptor territories on the periphery that are in functional correspondence with them. Examples of projection cortical zones are the area of \u200b\u200bthe posterior central gyrus already described in previous chapters (zone general types sensitivity) or the region of the spur groove located on the medial side of the occipital lobe (projection visual zone).
Association zones the cortex does not have direct connections with the periphery. They are located between the projection zones and have numerous associative links with these projection zones and with other associative zones. The function of the associative zones is to carry out a higher analysis and synthesis of many elementary and more complex components. Here, in essence, there is an understanding of the information entering the brain, the formation of ideas and concepts.
G.I. Polyakov in 1969, based on a comparison of the architectonics of the human cerebral cortex and some animals, found that associative
Rice. 14.6.Architectonic fields of the cerebral cortex [according to Brodman]. a - outer surface; b - medial surface.
zones in the human cerebral cortex are 50%, in the cortex of higher (humanoid) monkeys - 20%, in lower monkeys this figure is 10% (Fig. 14.7). Among the association areas of the cortex of the human brain, the same author suggested isolating secondary and tertiary fields. Secondary associative fields are adjacent to the projection ones. They carry out the analysis and synthesis of elementary sensations that still retains a specific orientation.
Tertiary associative fields are located mainly between the secondary ones and are overlapping zones of neighboring territories. They are related primarily to the analytical activity of the cortex, providing the highest mental functions inherent in man in their most complex intellectual and speech manifestations. Functional maturity of tertiary as-
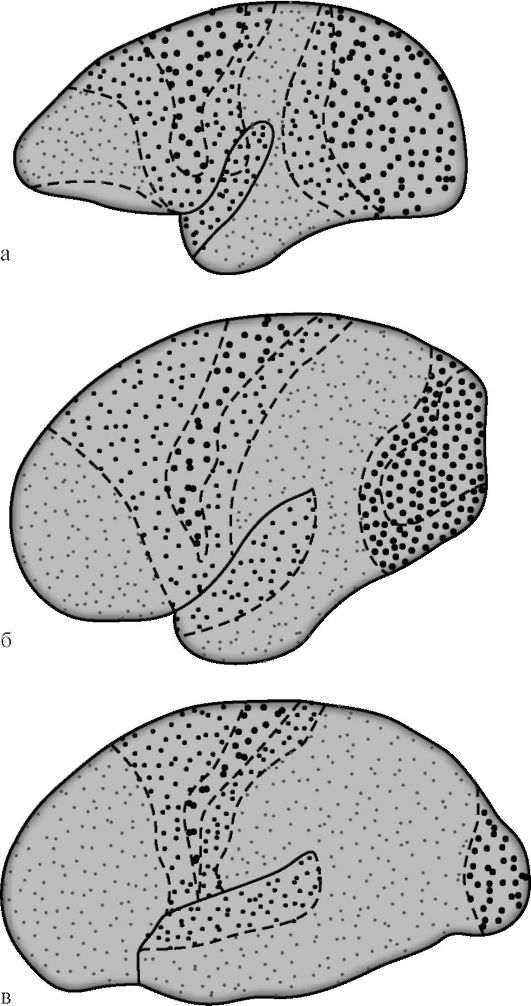
Rice. 14.7. Differentiation of projection and associative areas of the cerebral cortex during the evolution of primates [according to G.I. Polyakov]. a - the brain of the lower monkey; b - the brain of a higher ape; c - the human brain. Large dots indicate projection zones, small dots - associative ones. In lower monkeys, associative zones occupy 10% of the area of the cortex, in higher ones - 20%, in humans - 50%.
social fields of the cerebral cortex occurs most late and only in a favorable social environment. Unlike other cortical fields, the tertiary fields of the right and left hemispheres are characterized by a pronounced functional asymmetry.
14.8. TOPICAL DIAGNOSIS OF LESIONS OF THE BRAIN CORTEX
14.8.1. Manifestations of damage to the projection zones of the cerebral cortex
In the cortex of each cerebral hemisphere, behind the central gyrus, there are 6 projection zones.
1. In the anterior part of the parietal lobe, in the region of the posterior central gyrus (cytoarchitectonic fields 1, 2, 3) located projection zone of general types of sensitivity(Fig. 14.4). The areas of the cortex located here receive sensitive impulses coming along the projection pathways of general types of sensitivity from the receptor apparatus of the opposite half of the body. The higher the area of this projection zone of the cortex is, the lower the located parts of the opposite half of the body it has projection connections. Parts of the body with extensive reception (tongue, palmar surface of the hand) correspond to inadequately large parts of the area of projection zones, while other parts of the body (proximal limbs, torso) have a small area of cortical representation.
Irritation by the pathological process of the cortical zone of general types of sensitivity leads to an attack of paresthesia in parts of the body corresponding to the irritated areas of the cerebral cortex (sensitive Jacksonian seizure), which can turn into a secondary generalized paroxysm. The defeat of the cortical end of the analyzer of general types of sensitivity can cause the development of hypalgesia or anesthesia in the corresponding zone of the opposite half of the body, while the site of hypesthesia or anesthesia can be of a vertical circulatory or radicular-segmental type. In the first case, the sensitivity disorder manifests itself on the side opposite to the pathological focus in the region of the lips, thumb, or in the distal part of the limb with a circular border, sometimes like a sock or glove. In the second case, the zone of sensitivity disturbance has the form of a strip and is located along the inner or outer edge of the arm or leg; this is explained by the fact that the inner side of the limbs is presented in the anterior, and the outer side - in the posterior sections of the projection zone of the analyzer of general types of sensitivity.
2. Visual projection zone located in the cortex of the medial surface of the occipital lobe in the region of the spur groove (field 17). In this field, there is a stratification of the IV (internal granular) layer of the cortex with a bundle of myelin fibers into two sublayers. Separate sections of field 17 receive impulses from certain sections of the homonymous halves of the retinas of both eyes; while the impulses coming from the lower parts of the homonymous halves of the retinas reach
the cortex of the lower lip of the spur groove, and the impulses coming from the upper parts of the retinas are directed to the cortex of its upper lip.
The defeat of the pathological process of the visual projection zone leads to the appearance on the opposite side of a quadrant or complete homonymous hemianopia on the side opposite to the pathological focus. Bilateral damage to the cortical fields 17 or the projection visual pathways leading to them can lead to complete blindness. Irritation of the cortex of the visual projection zone can cause the appearance of visual hallucinations in the form of photopsies in the corresponding parts of the opposite halves of the visual fields.
3. Hearing projection area situated in the cortex of the convolutions of Heschl on the lower lip of the lateral (Sylvian) furrow (fields 41 and 42), which are, in fact, part of the superior temporal gyrus. Irritation of this zone of the cortex can cause the occurrence of auditory hallucinations (attacks of feeling noise, ringing, whistling, buzzing, etc.). The destruction of the auditory projection zone on the one hand can cause some hearing loss in both ears, to a greater extent on the opposite with respect to the pathological focus.
4 and 5. Olfactory and gustatory projection zones are on the medial surface of the vaulted gyrus (limbic region) of the brain. The first of them is located in the parahippocampal gyrus (field 28). The projection zone of taste is usually localized in the cortex of the opercular area (field 43). Irritation of the projection zones of smell and taste can cause their perversion or lead to the development of the corresponding olfactory and gustatory hallucinations. Unilateral loss of the function of the projection zones of smell and taste can cause a slight decrease in smell and taste, respectively, on both sides. Bilateral destruction of the cortical ends of the same analyzers is manifested by the absence of smell and taste on both sides, respectively.
6. Vestibular projection zone. Its localization is not specified. At the same time, it is known that the vestibular apparatus has numerous anatomical and functional connections. It is possible that the localization of the representation of the vestibular system in the cortex has not yet been clarified because it is polyfocal. N.S. Blagoveshchenskaya (1981) believes that in the cerebral cortex the vestibular projection zones are represented by several anatomical and functional interacting complexes, which are located in field 8, at the junction of the frontal, temporal and parietal lobes and in the zone of the central gyri, while it is assumed that each of these areas of the cortex performs its own functions. Field 8 is an arbitrary center of the gaze, its irritation causes the gaze to turn in the direction opposite to the pathological focus, changes in the rhythm and nature of the experimental nystagmus, especially soon after an epileptic seizure. In the cortex of the temporal lobe there are structures, the irritation of which causes dizziness, which manifests itself, in particular, in temporal lobe epilepsy; the defeat of the areas of representation of the vestibular structures in the cortex of the central gyri affects the state of the tone of the striated muscles. Clinical observations suggest that the nuclear-cortical vestibular pathways make a partial decussation.
It should be emphasized that signs of irritation of the listed projection zones can be a manifestation of the aura of an epileptic seizure corresponding in nature.
I.P. Pavlov considered it possible to consider the cortex of the precentral gyrus, which affects the motor functions and muscle tone of the predominantly opposite half of the body, with which it is connected primarily by the cortical-nuclear and cortical-spinal (pyramidal) pathways, as the projection zone of the so-called motor analyzer. This zone occupies first of all, field 4, on which the opposite half of the body is projected in an inverted form. This field contains the bulk of giant pyramidal cells (Betz cells), the axons of which make up 2-2.5% of all fibers of the pyramidal pathway, as well as medium and small pyramidal cells, which, together with the axons of the same cells located in the adjacent to the field 4 more extensive field 6, are involved in the implementation of monosynaptic and polysynaptic cortical-muscular connections. Monosynaptic connections provide mainly fast and precise targeted actions, depending on the contractions of individual striated muscles.
Damage to the lower parts of the motor zone usually leads to development on the opposite side brachiofacial (shoulder facial) syndrome or linguofaciobrachial syndrome, which are often observed in patients with impaired cerebral circulation in the basin of the middle cerebral artery, while combined paresis of the muscles of the face, tongue and arm, especially the shoulder in the central type.
Irritation of the cortex of the motor zone (fields 4 and 6) leads to the appearance of convulsions in the muscles projected onto this zone or muscle groups. More often, these are local convulsions of the type of Jacksonian epilepsy, which can transform into a secondary generalized epileptic seizure.
14.8.2. Manifestations of damage to the associative fields of the cerebral cortex
Between the projection zones of the cortex are association fields. They receive impulses mainly from the cells of the projection zones of the cortex. In the associative fields, the analysis and synthesis of information that has undergone primary processing in the projection fields takes place. Associative zones of the cortex of the upper parietal lobule provide a synthesis of elementary sensations, in connection with this, such complex types of sensitivity as a sense of localization, a sense of weight, a two-dimensional-spatial sense, as well as complex kinesthetic sensations are formed here.
In the region of the interparietal sulcus, there is an associative zone that provides a synthesis of sensations emanating from parts of one's own body. Damage to this region of the cortex leads to autopagnosia, those. to not recognizing or ignoring parts of one's own body, or to pseudomelia the feeling of having an extra arm or leg, and anosognosia - lack of awareness of a physical defect that has arisen in connection with the disease (for example, paralysis or paresis of a limb). Usually, all types of autopagnosia and anosognosia occur when the pathological process is located on the right.
The defeat of the lower parietal lobule can be manifested by a disorder in the synthesis of elementary sensations or the inability to compare the synthesized complex sensations with "there was once in the perception of similar
in the same way, on the basis of the results of which recognition occurs ”(V.M. Bekhterev). This is manifested by a violation of the two-dimensional spatial sense (grafoesthesia) and three-dimensional spatial sense (stereognosis) - astereognosis.
In the case of damage to the premotor zones of the frontal lobe (fields 6, 8, 44), frontal ataxia usually occurs, in which the synthesis of afferent impulses (kinesthetic afferentation) is disturbed, signaling the position of body parts in space that changes during the movements made.
In case of violation of the function of the cortex of the anterior parts of the frontal lobe, which has connections with the opposite hemisphere of the cerebellum (fronto-bridge-cerebellar connections), statokinetic disturbances occur on the opposite side of the pathological focus (frontal ataxia). Particularly distinct are violations of late developing forms of statokinetics - upright standing and upright walking. As a result, the patient has uncertainty, unsteadiness of gait. While walking, his body leans back. (Henner sign) he puts his feet in a straight line (fox gait) sometimes when walking there is a "braiding" of the legs. In some patients with damage to the anterior parts of the frontal lobes, a peculiar phenomenon develops: in the absence of paralysis and paresis and the preserved ability to move the legs in full, patients cannot stand (astasia) and walk (abasia).
The defeat of the associative zones of the cortex is often characterized by the development of clinical manifestations of a violation of higher mental functions (see Chapter 15).








