1. Pripomeň si z kurzu zoológie, ktoré časti mozgu majú všetky stavovce. Z akého oddelenia sú odvodené mozgové hemisféry? V ktorej skupine zvierat sa objavujú ako prvé?
Všetky stavovce sa vyznačujú prítomnosťou 5 častí mozgu: dreň zadný mozog, stredný mozog, diencephalon, predný mozog.
Všeobecný prehľad o štruktúre mozgových hemisfér
Nepochopenie mozgu koňa Nie je nezvyčajné počuť nasledujúce tvrdenia: V jednom koni sú dva kone: pravý kôň a spoľahlivý ľavý kôň a nepredvídateľný pravý kôň, čo sa týka dvoch hemisfér mozgu: ľavá hemisféra logika. Nikdy by ste nemali jazdiť na koni s použitím pravého mozgu a naučiť koňa najprv používať ľavý mozog. Kôň má dve vizuálne vedomia. Kôň nerozmýšľa, inštinktívne reaguje. Ale tieto tvrdenia nie sú založené na vedeckých dôkazoch.
Mozgové hemisféry sú deriváty predného mozgu a prvýkrát sa objavujú u obojživelníkov, ale sú slabo vyvinuté, mozgová kôra prakticky chýba.
2. Popíšte štruktúru hemisféry osoba. Nakreslite schematický rez mozgovými hemisférami, pričom na obrázku označte šedú hmotu kôry, šedú hmotu jadier, bielu hmotu a komory.
Pohľad na históriu evolúcie cicavcov na hominidov a miesto koňa v tejto histórii je spôsob, ako argumentovať proti týmto chybám v analýze správania koní. Trieda cicavcov má niekoľko oddelení alebo rádov, ako sú hlodavce, mäsožravce, hmyzožravce, kopytníky a primáty.
Cicavce sú zvieratá charakterizované produkciou mlieka z mliečnych žliaz samice. Kôň patrí do triedy cicavcov a do radu kopytníkov, pretože okraj prstov je pokrytý kopytom a končatiny spočívajú na poslednej falange. Kôň bol však domestikovaný až neskôr, prvé stopy domestikácie sa datujú od 500 do tisíc rokov. Evolúcia mozgu u niektorých dospelých cicavcov. celková hmotnosť mozgu v gramoch.
U ľudí je predný mozog reprezentovaný dvoma hemisférami a corpus callosum spájajúcim hemisféry. Veľké hemisféry: pravá a ľavá - pokrývajú stredný a diencefalón a tvoria až 80% hmoty mozgu dospelého človeka. Na povrchu každej hemisféry je veľa brázd a zákrut. Existujú 4 hlavné sulci (centrálny, laterálny a parietálno-okcipitálny), ktoré rozdeľujú každú hemisféru na laloky. Povrchovú vrstvu tvorí sivá hmota (kôra), pod ňou je biela hmota, pozostávajúca z axónov nervových buniek, ktorých telá ležia v kôre alebo ktoré prenášajú informácie do buniek kôry. Veľké nahromadenia šedej hmoty (subkortikálne jadrá) a dutín (laterálne komory) ležia v hrúbke bielej hmoty. Cez komory mozgu a centrálny kanál miecha cirkuluje mozgovomiechový mok, ktorý zabezpečuje výživu podkôrových štruktúr.
Celková hmotnosť mozgu vyjadrená ako percento telesnej hmotnosti. Primáty a hominidy. V rámci triedy cicavcov existuje aj rad primátov, ktoré sa objavujú na začiatku treťohôr. Medzi týmito primátmi sú pongidy alebo antropopodné ľudoopy a hominidy. Hominid je rodina bipedálnych primátov, t.j. austratopitov ako Lucy a neskôr samcov.
Evolúcia mozgu Štúdium evolúcie živých vecí v priebehu miliónov rokov od prvých cicavcov po ľudí umožnilo pozorovať všeobecný trend mozgu k zvýšeniu objemu globálne av prospech špecifických oblastí mozgu. U cicavcov bol tento nárast najmä v prospech mozgových hemisfér a mozočku, takže na rozdiel od vtákov a plazov sa len u cicavcov vyvinula neorktéria so 6 bunkovými vrstvami, tento nocort predstavuje viac ako polovicu objemu mozgu u najvyspelejších cicavcov.
3. Čo je to mozgová kôra? Kde to je?
Povrchová vrstva šedej hmoty mozgových hemisfér sa nazýva kôra. Kôra pozostáva z niekoľkých vrstiev telies neurónov, ktoré sa líšia štruktúrou a funkciou. Predpokladá sa, že jeho zloženie obsahuje asi 12-18 miliárd buniek, hrúbka je 1,5-4,5 mm a plocha je 1,7-2,5 tisíc cm2.
Štruktúra predného laloku
Evolúcia hominidov tiež zaznamenala zvýšenie lebečnej kapacity prostredníctvom postupného rastu mozgu a mozočka. Jedna z vlastností ľudský mozog na rozdiel od iných cicavcov je jeho anatomická a funkčná asymetria. Napríklad veľké časti ľavej hemisféry sa špecializujú na tvorbu a porozumenie jazyka, na písanie a čítanie. Pravá hemisféra je špecifickejšie spojená s priestorovou orientáciou pri detekcii vizuálne formy a hudobné znalosti. Ľavá hemisféra je dominantnejšia pre procesy, logická, analytická a pravá pre holistické procesy.
4. Vysvetlite význam brázd a záhybov na povrchu mozgových hemisfér.
Brázdy a gyrus výrazne zväčšujú povrch mozgovej kôry, podľa niektorých zdrojov až 10-12 krát.
5. Akú funkciu má biela hmota mozgových hemisfér?
Biela hmota tvorí cesty spájajúce kôru a kôru so zvyškom nervového systému.
Asymetria mozgu teda je vlastnosťčloveka, rovnako ako nedostatok pozoruhodnej siete u opíc, ktoré stále majú symetrické mozgy, staroveký norcort si zachováva svoje symetrické fungovanie, takže logika by sa nemala naznačovať. u koňa je jedna z dvoch hemisfér v určitej funkcii, hovoria niektorí noví majstri.
Ale ani u ľudí nie je mozgová hemisféra nikdy jediná, ktorá zasahuje do funkcie, ale zdá sa, že je len viac prepojená, implicitne v tejto funkcii. V skutočnosti zostávajú základné funkcie mozgu do značnej miery distribuované vo všetkých dvoch hemisférach mozgu. V tejto súvislosti je tiež dôležité pripomenúť prítomnosť štruktúry corpus callosum u ľudí aj u koní. Táto štruktúra je skutočné spojenie medzi dvoma hemisférami, ktoré spolu komunikujú. Tvrdiť teda, že spojenie medzi hemisférami mozgu na koni je absolútne nesprávne.
6. Aké laloky sú izolované v mozgových hemisférach?
V hemisférach sa rozlišujú predné, 2 parietálne, 2 temporálne a okcipitálne laloky
7. Rozlišujte medzi pojmami "laloky mozgových hemisfér" a "zóny mozgových hemisfér." Uveďte príklady, keď sa zhodujú, nezhodujú sa.
Laloky mozgových hemisfér sú rozdelenie povrchu kôry podľa anatomického princípu: v každej hemisfére sa rozlišujú čelné, okcipitálne, parietálne a temporálne laloky.
Kortikálne zóny - časť mozgovej kôry, charakterizovaná jednotnosťou štruktúry a vykonávaných funkcií.
V našom mozgu sa laloky a zóny nezhodujú, jeden lalok sa skladá z niekoľkých zón. Okcipitálny lalok pozostáva z oblastí videnia a vizuálneho rozpoznávania. V temporálnom laloku a vedľa neho sú zóny čuchu, sluchu a chuti.
8. Pri prehliadke nevidomého pacienta sa zistilo, že jeho oči a zrakové nervy nie sú poškodené. Prečo nevidí?
Neschopnosť vidieť môže byť spôsobená okrem poškodenia zraku alebo zrakových nervov aj poškodením zrakových zón okcipitálnych lalokov mozgových hemisfér.
9. Pomocou ďalších zdrojov informácií zistite, či sa líšia funkcie ľavej a pravej hemisféry mozgu.
Medzi pravou a ľavou hemisférou mozgu existuje „funkčná asymetria“, to znamená, že ich funkcie sú odlišné. Preukázalo sa to pri pokusoch o prerezávaní komunikačných liniek medzi hemisférami (následne sa takéto operácie začali z medicínskych dôvodov vykonávať pri niektorých chorobách, ako je Parkinsonova choroba). Praváci majú ľavú hemisféru, zatiaľ čo ľaváci majú pravú hemisféru. Za to je zodpovedná pravá hemisféra kreatívne myslenie, tvorí základ kreativity, robí neštandardné rozhodnutia; Vizuálna oblasť pravej hemisféry je zodpovedná za rozpoznávanie tváre. Ľavá hemisféra poskytuje logické uvažovanie a abstraktné myslenie, obsahuje centrá ústnej a písomnej reči, formovanie rozhodnutí; zraková zóna je zodpovedná za rozpoznávanie písmen a číslic. Preto, žartom, praváci sa nazývajú matematici a ľaváci sú umelci.
10. Existujú každodenné pojmy „mužská logika“ a „ženská logika“. Existujú nejaké dôvody na takéto rozdiely?
Vo vonkajšej a vnútornej štruktúre nie je rozdiel medzi mozgom mužov a žien, ale napriek tomu, že štruktúra nášho mozgu je takmer rovnaká, sme individuálni. Hlavné rozdiely vo vnímaní sveta podľa „mužského“ alebo „ženského“ typu sú uvedené v rané detstvo v závislosti od našej výchovy a pod vplyvom hormónov pohlavných žliaz počas dospievania, ale všetky sú len psychického charakteru.
14.1. VŠEOBECNÉ USTANOVENIA
Koncový mozog (telencephalon) alebo veľký mozog (cerebrum), nachádza sa v supratentoriálnom priestore lebečnej dutiny pozostáva z dvoch veľkých
hemisféry (gemispherium cerebralis),oddelené hlbokou pozdĺžnou štrbinou (fissura longitudinis cerebri), v ktorom je ponorený polmesiac mozgu (falx cerebri) predstavujúce duplikáciu dura mater. Veľké hemisféry mozgu tvoria 78 % jeho hmoty. Každá z mozgových hemisfér má laloky: čelné, parietálne, temporálne, okcipitálne a limbické. Pokrývajú štruktúry diencefala a mozgového kmeňa a mozočka umiestnené pod plášťom mozočka (subtentoriálne).
Každá z mozgových hemisfér má tri povrchy: horná laterálna, alebo konvexitná (obr. 14.1a), - konvexná, smerujúca ku kostiam lebečnej klenby; vnútorný (obr. 14.1b), priliehajúci k veľkému falciformnému výbežku a spodný, čiže bazálny (obr. 14.1c), opakujúci reliéf spodiny lebečnej (predné a stredné jamky) a cerebelárneho čapu. V každej hemisfére sa rozlišujú tri okraje: horný, spodný vnútorný a spodný vonkajší a tri póly: predný (čelný), zadný (okcipitálny) a laterálny (temporálny).
Dutina každej mozgovej hemisféry je bočná komora mozgu zatiaľ čo ľavá bočná komora je rozpoznaná ako prvá, pravá - druhá. Bočná komora má centrálnu časť umiestnenú hlboko v parietálnom laloku (lobus parietalis) a tri rohy vystupujúce z neho: predný roh preniká do čelného laloku (lobus frontalis), nižšia - do časovej (lobus temporalis), zadné - v okcipitálnom (lobus occipitalis). Každá z bočných komôr komunikuje s treťou komorou mozgu cez interventrikulárnu komoru diera Monroe.
Centrálne úseky mediálneho povrchu oboch hemisfér sú vzájomne prepojené mozgovými komisúrami, z ktorých najmasívnejším je corpus callosum, a štruktúrami diencephala.
Telencephalon, podobne ako ostatné časti mozgu, pozostáva zo šedej a bielej hmoty. Sivá hmota sa nachádza v hĺbke každej hemisféry, tvorí tam podkôrové uzliny a pozdĺž okraja voľných plôch hemisféry, kde tvorí mozgovú kôru.
Hlavné otázky súvisiace so stavbou, funkciami bazálnych ganglií a variantmi klinického obrazu pri ich postihnutí sú rozobraté v kapitolách 5, 6. Mozgová kôra je približne
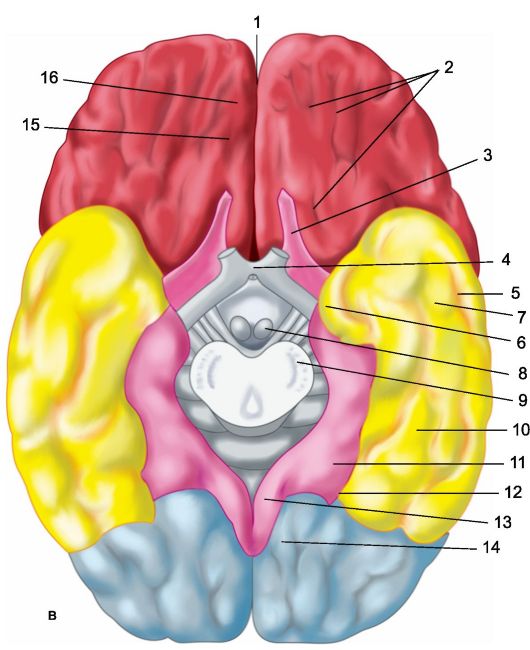
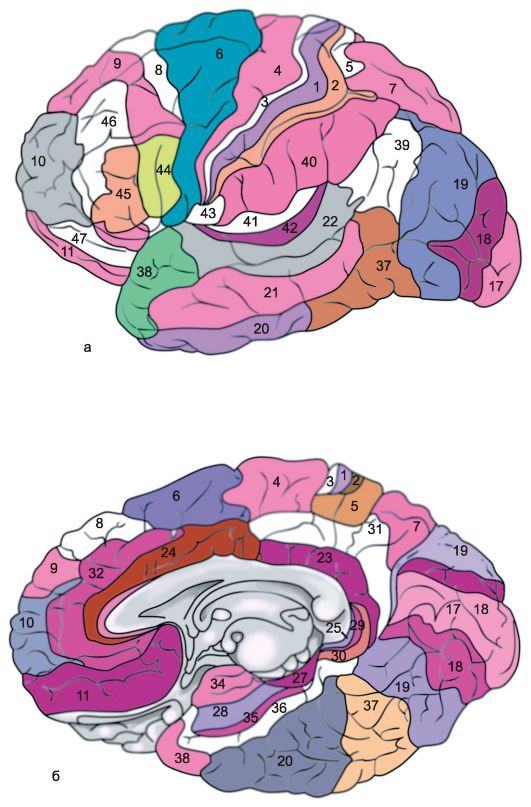
Ryža. 14.1.Hemisféry mozgu.
a - horný bočný povrch ľavej hemisféry: 1 - centrálny sulcus; 2 - orbitálna časť dolného čelného gyru; I - čelný lalok; 3 - precentrálny gyrus; 4 - predcentrálna brázda; 5 - horný frontálny gyrus; 6 - stredný čelný gyrus; 7 - tegmentálna časť dolného čelného gyru; 8 - dolný čelný gyrus; 9 - bočná brázda; II - parietálny lalok: 10 - postcentrálny gyrus; 11 - postcentrálna brázda; 12 - intraparietálna drážka; 13 - supramarginálny gyrus; 14 - uhlový gyrus; III - temporálny lalok: 15 - horný temporálny gyrus; 16 - horný temporálny sulcus; 17 - stredný temporálny gyrus; 18 - stredný temporálny sulcus; 19 - dolný temporálny gyrus; IV - okcipitálny lalok: b - mediálny povrch pravej hemisféry: 1 - paracentrálny lalok, 2 - precuneus; 3 - parieto-okcipitálny sulcus; 4 - klin, 5 - lingválny gyrus; 6 - laterálny okcipitotemporálny gyrus; 7 - parahippokampálny gyrus; 8 - hák; 9 - klenba; 10 - corpus callosum; 11 - horný frontálny gyrus; 12 - cingulárny gyrus; c - spodný povrch mozgových hemisfér: 1 - pozdĺžna interhemisferická trhlina; 2 - orbitálne brázdy; 3 - čuchový nerv; 4 - optický chiasma; 5 - stredný temporálny sulcus; 6 - hák; 7 - dolný temporálny gyrus; 8 - mastoidné telo; 9 - základňa mozgového kmeňa; 10 - laterálny okcipitotemporálny gyrus; 11 - parahippokampálny gyrus; 12 - vedľajšia drážka; 13 - cingulárny gyrus; 14 - lingválny gyrus; 15 - čuchová drážka; 16 - priamy gyrus.
3-násobok povrchu hemisfér viditeľných pri externom vyšetrení. Je to spôsobené tým, že povrch mozgových hemisfér je zložený, má početné priehlbiny - brázdy (sulci cerebri) a nachádza sa medzi nimi konvolúcie (gyri cerebri). Mozgová kôra pokrýva celý povrch záhybov a brázd (odtiaľ jej iný názov je pálium - plášť), pričom niekedy preniká do veľkej hĺbky až do hmoty mozgu.
Závažnosť a umiestnenie brázd a konvolúcií mozgových hemisfér sú do určitej miery variabilné, ale hlavné sa tvoria v procese ontogenézy a sú konštantné, charakteristické pre každý normálne vyvinutý mozog.
14.2. HLAVNÉ DRÁHY A ÚCHYTKY HEMISfér MOZGU
Horný bočný (konvexitný) povrch hemisfér (obr. 14.1a). Najväčší a najhlbší bočné brázda (sulcus lateralis),alebo sylvian brázda, - oddeľuje prednú a prednú časť parietálneho laloku od temporálneho laloku umiestneného nižšie. Čelný a parietálny lalok sú oddelené centrálna, alebo Rolandova brázda(sulcus centralis), ktorá pretína horný okraj pologule a ide dole a dopredu pozdĺž jej konvexného povrchu, mierne kratšia od bočnej drážky. Parietálny lalok je oddelený od okcipitálneho laloku, ktorý sa nachádza za ním, parietálno-okcipitálnymi a priečnymi okcipitálnymi drážkami prechádzajúcimi pozdĺž stredného povrchu hemisféry.
V prednom laloku pred centrálnym gyrusom a rovnobežne s ním je precentrálny (gyrus precentralis), alebo predná centrálna, gyrus, ktorý je vpredu ohraničený precentrálnym sulkusom (sulcus precentralis). Horné a dolné frontálne drážky odchádzajú dopredu od precentrálneho sulcus a rozdeľujú konvexný povrch predných častí frontálneho laloku na tri frontálne gyrus - horný, stredný a dolný (gyri frontales superior, media et inferior).
Predná časť konvexitného povrchu parietálneho laloku sa nachádza za centrálnym sulcus postcentral (gyrus postcentralis), alebo zadný centrálny, gyrus. Za ním je ohraničený postcentrálnym sulcusom, z ktorého sa dozadu tiahne intraparietálny sulcus. (sulcus intraparietalis), oddeľujúce horný a dolný parietálny lalok (lobuli parietales superior et inferior). V dolnom parietálnom laloku sa zasa rozlišuje supramarginálny gyrus (gyrus supramarginalis), obklopujúce zadnú časť laterálnej (Sylviovej) drážky a uhlový gyrus (girus angularis), ohraničujúce zadnú časť horného temporálneho gyru.
Na konvexitnom povrchu okcipitálneho laloku mozgu sú brázdy plytké a môžu sa výrazne líšiť, v dôsledku čoho je tiež variabilný charakter medzi nimi umiestnených zákrutov.
Konvexitná plocha spánkového laloka je rozdelená horným a dolným temporálnym žliabkom, ktoré sú takmer paralelné s laterálnym (Sylviovským) žliabkom a rozdeľujú konvexnú plochu spánkového laloku na horný, stredný a dolný temporálny gyri. (gyri temporales superior, media et inferior). Gyrus temporalis superior tvorí dolný pysk laterálneho (Sylviovho) sulcus. Na svojom povrchu obrátený
na strane bočnej brázdy je niekoľko priečnych malých brázd, ktoré zvýrazňujú malý priečny gyrus (gyrus z Geschlu), ktoré možno vidieť len roztiahnutím okrajov bočnej brázdy.
Predná časť laterálnej (Sylvian) ryhy je priehlbina so širokým dnom, tvoriaca tzv. ostrov (ostrov) alebo ostrovný lalok (lubus insularis). Horný okraj bočnej brázdy pokrývajúcej tento ostrov je tzv pneumatika (operculum).
Vnútorný (mediálny) povrch hemisféry (obr. 14.1b). Centrálna časť vnútorného povrchu hemisféry je úzko spojená so štruktúrami diencephala, z ktorých je ohraničená tými, ktoré súvisia s veľkým mozgom. trezor (fornix) a corpus callosum (corpus callosum). Ten je zvonku ohraničený brázdou corpus callosum (sulcus corporis callosi), počnúc jeho prednou časťou - zobákom (reštaurácia) a končí na jeho zosilnenom zadnom konci (slezina). Tu sulcus corpus callosum prechádza do hlbokej hipokampálnej ryhy (sulcus hippocampi), ktorá preniká hlboko do hmoty hemisféry, vtláča ju do dutiny dolného rohu laterálnej komory, v dôsledku čoho dochádza k tzv. -vytvára sa takzvaná amónna rohovina.
Mierne odstupujúce od sulcus corpus callosum a hippocampal sulcus sa nachádzajú corpus callosum, subparietálne a nazálne sulci, ktoré sú vzájomným pokračovaním. Tieto drážky ohraničujú z vonkajšej strany oblúkovitú časť mediálneho povrchu mozgovej hemisféry, tzv. limbický lalok(lobus limbicus). V limbickom laloku sú dve konvolúcie. Horná časť limbického laloku je horný limbický (superior marginálny) alebo pletenec, gyrus (girus cinguli), spodnú časť tvorí gyrus limbický alebo gyrus morského koníka (girus hippocampi), alebo parahipokampálny gyrus (girus parahypocampalis), pred ktorým je háčik (uncus).
Okolo limbického laloku mozgu sú útvary vnútorného povrchu čelného, parietálneho, okcipitálneho a temporálneho laloku. Väčšinu vnútorného povrchu predného laloku zaberá stredná strana horného frontálneho gyru. Na hranici medzi čelným a parietálnym lalokom mozgovej hemisféry sa nachádza paracentrálny lalok (lobulis paracentralis), ktorý je akoby pokračovaním predného a zadného centrálneho gyru na mediálnom povrchu hemisféry. Na hranici medzi parietálnym a okcipitálnym lalokom je jasne viditeľný parietálno-okcipitálny sulcus. (sulcus parietooccipitalis). Zo spodnej časti odchádza späť ostrohová brázda (sulcus calcarinus). Medzi týmito hlbokými brázdami je trojuholníkový gyrus, známy ako klin. (cuneus). Pred klinom je štvoruholníkový gyrus, súvisiaci s parietálnym lalokom mozgu, precuneus.
Spodný povrch hemisféry (obr. 14.1c). Spodný povrch mozgovej hemisféry pozostáva z formácií čelných, temporálnych a okcipitálnych lalokov. Časť predného laloku susediaca so stredovou čiarou je priamy gyrus (girus rectus). Vonku je ohraničený čuchovým žliabkom (sulcus olfactorius), ku ktorým zospodu priliehajú útvary čuchového analyzátora: čuchový bulbus a čuchový trakt. Bočne od nej, až po laterálnu (Sylvovu) drážku, ktorá sa tiahne k spodnej ploche predného laloku, sú malé orbitálne gyri (gyri orbitalis). Bočné časti spodného povrchu hemisféry za laterálnym sulcusom sú obsadené dolným temporálnym gyrusom. Mediálne k nemu je laterálny temporo-okcipitálny gyrus. (gyrus occipitotemporalis lateralis), alebo vretenovitá drážka. Pred-
jeho vnútorné časti hraničia s gyrus hipokampu a zadné - s lingválnym (gyrus lingualis) alebo mediálny temporoccipitálny gyrus (gyrus occipitotemporalis medialis). Ten so svojim zadným koncom prilieha k ostrohovej drážke. Predné časti fusiformných a jazykových gyri patria k temporálnemu laloku a zadné časti k okcipitálnemu laloku mozgu.
14.3. BIELA HMOTA VEĽKÝCH POLOGULÍ
Biela hmota mozgových hemisfér pozostáva z nervových vlákien, hlavne myelínu, ktoré tvoria dráhy, ktoré zabezpečujú spojenie medzi neurónmi kôry a zhlukmi neurónov, ktoré tvoria talamus, subkortikálne uzliny a jadrá. Hlavná časť bielej hmoty mozgových hemisfér sa nachádza v jej hĺbke polooválny stred, alebo žiarivá koruna (corona radiata), pozostávajúce hlavne z aferentných a eferentných projekcia dráhy spájajúce mozgovú kôru so subkortikálnymi uzlinami, jadrami a retikulárnou substanciou diencefala a mozgového kmeňa, so segmentmi miechy. Sú obzvlášť kompaktne umiestnené medzi talamom a subkortikálnymi uzlinami, kde tvoria vnútorné puzdro opísané v kapitole 3.
Nervové vlákna, ktoré spájajú časti kôry jednej hemisféry, sa nazývajú asociatívne. Čím kratšie sú tieto vlákna a spojenia, ktoré vytvárajú, tým sú povrchnejšie; dlhšie asociatívne spojenia, umiestnené hlbšie, spájajú relatívne vzdialené časti mozgovej kôry (obr. 14.2 a 14.3).
Vlákna, ktoré spájajú mozgové hemisféry, a preto majú spoločnú priečnu orientáciu, sa nazývajú komisurálny, alebo spánok. Komisurálne vlákna spájajú identické časti mozgových hemisfér, čím vytvárajú možnosť kombinácie ich funkcií. Tvoria sa tri hroty veľký mozog: najmasívnejší z nich - corpus callosum (corpus callosum), okrem toho tvoria komisurálne vlákna predná komisúra, nachádza sa pod zobákom corpus callosum (rostrum corporis collosum) a prepojenie oboch čuchových oblastí, ako aj komisúra klenby (commissura fornicis), alebo hipokampálna komisura tvorená vláknami spájajúcimi štruktúry amonných rohov oboch hemisfér.
V prednej časti corpus callosum sú vlákna spájajúce čelné laloky, ďalej vlákna spájajúce parietálny a spánkový lalok, zadná časť corpus callosum spája okcipitálne laloky mozgu. Predná komizúra a komizúra fornixu zjednocujú hlavne úseky starej a starej kôry oboch hemisfér, predná komizúra navyše poskytuje spojenie medzi ich strednými a dolnými temporálnymi gyri.
14.4. Čuchový systém
V procese fylogenézy je vývoj veľkého mozgu spojený s formovaním čuchového systému, ktorého funkcie prispievajú k zachovaniu životaschopnosti zvierat a majú nemalý význam pre život človeka.
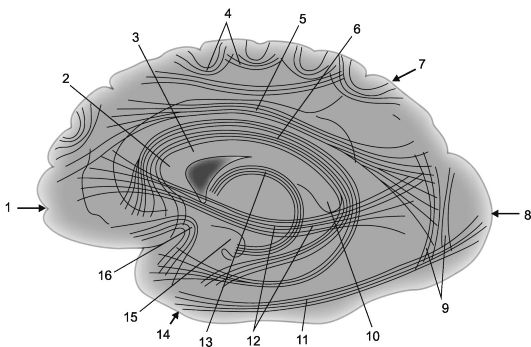
Ryža. 14.2.Asociatívne kortikálno-kortikálne spojenia v mozgových hemisférach [podľa V.P. Vorobyov].
1 - čelný lalok; 2 - koleno corpus callosum; 3 - corpus callosum; 4 - oblúkové vlákna; 5 - horný pozdĺžny nosník; 6 - cingulárny gyrus; 7 - parietálny lalok, 8 - okcipitálny lalok; 9 - vertikálne zväzky Wernickeho; 10 - valček corpus callosum;
11 - spodný pozdĺžny nosník; 12 - subkauzálny zväzok (frontálny-okcipitálny spodný zväzok); 13 - klenba; 14 - temporálny lalok; 15 - hák gyrus hipokampu; 16 - zväzky háčikov (fasciculus uncinatus).
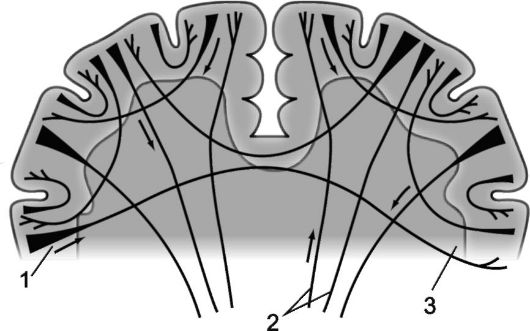
Ryža. 14.3.Myeloarchitektonika mozgových hemisfér.
1 - projekčné vlákna; 2 - komisurálne vlákna; 3 - asociatívne vlákna.
14.4.1. Štruktúra čuchového systému
Telá prvých neurónov čuchového systému sa nachádzajú v sliznici hlavne nos horná časť nosovej priehradky a horný nosový priechod. Čuchové bunky sú bipolárne. Ich dendrity vystupujú na povrch sliznice a končia tu špecifickými receptormi a axóny sú zoskupené v tzv čuchové vlákna (filiolfactorii), ktorých počet na každej strane je asi dvadsať. Takéto zväzok čuchových vlákien a tvorí I kraniálny, čiže čuchový nerv(obr. 14.4). Tieto vlákna prechádzajú do prednej (čuchovej, čuchovej) lebečnej jamky cez etmoidnú kosť a končia v bunky, ktoré sa tu nachádzajú čuchové žiarovky. Čuchové bulby a proximálne čuchové dráhy sú v skutočnosti dôsledkom výbežkov substancie veľkého mozgu vzniknutých v procese ontogenézy a predstavujú s ňou súvisiace štruktúry.
Čuchové žiarovky obsahujú bunky, ktoré sú telami druhých neurónov. čuchová dráha, ktorej axóny tvoria čuchové ústrojenstvo (tracti olfactorii), umiestnené pod čuchovými drážkami, laterálne k priamym zákrutom umiestneným na bazálnej ploche čelných lalokov. Čuchové dráhy smerujú dozadu do subkortikálnych čuchových centier. Približujúc sa k prednej perforovanej platni sú vlákna čuchového traktu rozdelené na mediálne a laterálne zväzky, ktoré na každej strane tvoria čuchový trojuholník. Neskôr sú tieto vlákna vhodné k telám tretích neurónov čuchového analyzátora, ktoré sa nachádzajú

Ryža. 14.4.Čuchový analyzátor.
1 - čuchové bunky; 2 - čuchové vlákna (spolu tvoria čuchové nervy); 3 - čuchové žiarovky; 4 - čuchové cesty; 5 - čuchové trojuholníky; 6 - parahippokampálny gyrus; 7 - projekčná zóna čuchového analyzátora (zjednodušená schéma).
v perialmdlových a subkalóznych oblastiach, v jadrách priehľadnej priehradky, umiestnenej pred prednou komisurou. Predná komisura spája obe čuchové oblasti a tiež zabezpečuje ich spojenie s limbickým systémom mozgu. Časť axónov tretích neurónov čuchového analyzátora prechádzajúca cez prednú komisuru mozgu sa prekríži.
Axóny tretích neurónov analyzátor čuchu umiestnený v podkôrových čuchových centrách, smerujúci k fylogeneticky stará kôra mediobazálna plocha spánkového laloka (k piriformnému a parahipokampálnemu gyru a k háčiku), kde sa nachádza projekčná čuchová zóna, alebo kortikálny koniec čuchového analyzátora (pole 28, podľa Brodmanna).
Čuchový systém je tak jediným zmyslovým systémom, v ktorom špecifické impulzy obchádzajú talamus na ceste z receptorov do kôry. však Čuchový systém má obzvlášť výrazné spojenie s limbickými štruktúrami mozgu a prostredníctvom neho prijímané informácie majú významný vplyv na stav emocionálnej sféry a funkcie autonómneho nervového systému. Vône môžu byť príjemné a nepríjemné, ovplyvňujú chuť do jedla, náladu, môžu vyvolať rôzne vegetatívne reakcie, najmä nevoľnosť, vracanie.
14.4.2. Vyšetrovanie čuchu a významu jeho porúch pre lokálnu diagnostiku
Pri vyšetrovaní čuchového stavu je potrebné zistiť, či pacient zapácha, či sú tieto vnemy na oboch stranách rovnaké, či pacient rozlišuje charakter pociťovaných pachov, či má čuchové halucinácie – záchvatové vnemy, ktoré v prostredí chýbajú.
Na štúdium čuchu sa používajú pachové látky, ktorých pach nie je ostrý (štipľavé pachy môžu spôsobiť podráždenie receptorov trojklaného nervu nachádzajúcich sa v nosovej sliznici) a sú pacientovi známe (inak je ťažké rozpoznať perverzia čuchu). Čuch sa kontroluje na každej strane zvlášť, pričom druhá nosová dierka musí byť uzavretá. Môžete použiť špeciálne pripravené sady slabých roztokov pachových látok (mäta, decht, gáfor atď.), V praktickej práci možno použiť aj improvizované prostriedky (ražný chlieb, mydlo, banán atď.).
Znížený čuch - hyposmia, nedostatok vône - anosmia, zvýšený čuch - hyperosmia, perverzia pachov dysosmia, pocit pachu pri absencii podnetu - parosmia, subjektívny pocit nepríjemného zápachu, ktorý skutočne existuje a je spôsobený organickou patológiou v nosohltane - Kakosmiya, v skutočnosti neexistujúce pachy, ktoré pacient pociťuje záchvatovito – čuchové halucinácie – sú častejšie čuchovou aurou epilepsie temporálneho laloka, čo môže byť spôsobené rôzne dôvody, najmä nádor spánkového laloku.
Hyposmia alebo anosmia na oboch stranách je zvyčajne výsledkom poškodenia nosovej sliznice v dôsledku akútneho kataru, chrípky, alergickej rinitídy, atrofie sliznice
nos v dôsledku chronickej rinitídy a dlhodobého používania vazokonstrikčných nosových kvapiek. Chronická rinitída s atrofiou nosovej sliznice (atrofická rinitída), Sjögrenova choroba odsudzuje človeka na pretrvávajúcu anosmiu. Bilaterálna hyposmia môže byť spôsobená hypotyreózou, diabetes mellitus, hypogonadizmom, zlyhaním obličiek, dlhodobým kontaktom s ťažkými kovmi, formaldehydom atď.
však jednostranná hyposmia alebo anosmia je často výsledkom intrakraniálneho nádoru, častejšie meningiómu prednej lebečnej (čuchovej) jamky, ktorý tvorí až 10 % intrakraniálnych meningiómov, ako aj niektoré gliové nádory čelového laloku. Poruchy čuchu sa vyskytujú v dôsledku stlačenia čuchového traktu na strane patologického zamerania a môžu byť po určitú dobu jediným ohniskovým príznakom ochorenia. Nádory je možné zobraziť pomocou CT alebo MRI skenovania. S nárastom meningiómu čuchovej jamky sa spravidla rozvíjajú duševné poruchy charakteristické pre frontálny syndróm (pozri kapitolu 15).
Jednostranné poškodenie častí čuchového analyzátora umiestnených nad jeho subkortikálnymi centrami v dôsledku neúplného prehĺbenia dráh na úrovni prednej cerebrálnej komizúry zvyčajne nevedie k výraznému zníženiu čuchu. Podráždenie patologickým procesom kôry mediobazálnych častí spánkového laloku, predovšetkým gyrus parahipocampal a jeho hák, môže spôsobiť záchvatovitý výskyt čuchové halucinácie. Pacient náhle začne bezdôvodne zapáchať, často nepríjemnej povahy (zápach spáleniny, zhnitého, zhnitého, spáleného atď.). Čuchové halucinácie v prítomnosti epileptogénneho zamerania v mediobazálnych oblastiach spánkového laloku mozgu môže byť prejavom aury epileptického záchvatu. Poškodenie proximálnej časti, najmä kortikálneho konca čuchového analyzátora, môže spôsobiť miernu bilaterálnu (viac na opačnej strane) hyposmiu a zhoršenú schopnosť identifikovať a rozlišovať pachy (čuchová agnózia). Posledná forma poruchy čuchu, ktorá sa prejavuje v starobe, je s najväčšou pravdepodobnosťou spojená s porušením funkcie kôry v dôsledku atrofických procesov v jej projekčnej čuchovej zóne.
14.5. LIMBIC-RETIKULÁRNY KOMPLEX
V roku 1878 P. Broca(Broca P., 1824-1880) pod názvom "veľký okrajový alebo limbický lalok" (z lat. limbus - okraj) spájal hipokampus a gyrus cingulate, vzájomne prepojené pomocou isthmu cingulate gyrus, umiestneného nad hrebeňom corpus callosum.
V roku 1937 D. Papets(Papez J.) na základe experimentálnych údajov vzniesol odôvodnenú námietku voči predtým existujúcemu konceptu zapojenia mediobazálnych štruktúr mozgových hemisfér najmä do poskytovania čuchu. On navrhol, že hlavná časť mediobazálnych častí mozgovej hemisféry, vtedy nazývanej čuchový mozog (rhinencephalon), ku ktorému patrí limbický lalok, je morfologickým základom nervového mechanizmu afektívneho správania a spojil ich pod názvom"emocionálny kruh" ktorý zahŕňal hypotalamus,
predné jadrá talamu, gyrus cingulate, hippocampus a ich spojenia. Odvtedy tieto štruktúry fyziológovia označujú aj ako okolo Papeza.
koncepcie "viscerálny mozog" navrhol P.D. McLean (1949), teda označuje komplexnú anatomicko-fyziologickú asociáciu, ktorá sa od roku 1952 nazýva tzv. „limbický systém“. Neskôr sa ukázalo, že limbický systém sa podieľa na výkone rôznych funkcií a v súčasnosti sa väčšina z nich, vrátane cingulárneho a hipokampálneho (parahipokampálneho) gyru, zvyčajne spája do limbickej oblasti, ktorá má početné spojenia so štruktúrami retikulárna formácia, tvoriaca s ňou limbicko-retikulárny komplex široký kruh fyziologické a psychické procesy.
Aktuálne do limbický lalok je zvykom pripisovať prvky starej kôry (archiokortex), pokrývajúcej gyrus dentatus a gyrus hippocampu; starodávna kôra (paleokortex) predného hipokampu; ako aj stredná alebo intermediárna kôra (mezokortex) gyrus cingulate. Termín "limbický systém" zahŕňa komponenty limbického laloku a súvisiace štruktúry – entorinálne (zaberajúce väčšinu parahipokampálneho gyru) a septálne oblasti, ako aj komplex amygdaly a mastoidné telo (Duus P., 1995).
Mastoidné telo spája štruktúry tohto systému so stredným mozgom a s retikulárnou formáciou. Impulzy pochádzajúce z limbického systému sa môžu prenášať cez predné jadro talamu do gyrus cingulate a do neokortexu po dráhach tvorených asociatívnymi vláknami. Impulzy pochádzajúce z hypotalamu môžu dosiahnuť orbitofrontálny kortex a mediálne dorzálne jadro talamu.
Početné priame a spätná väzba zabezpečujú prepojenie a vzájomnú závislosť limbických štruktúr a mnohých útvarov diencefala a ústnej časti mozgového kmeňa (nešpecifické jadrá talamu, hypotalamu, putamenu, uzdičky, retikulárna formácia mozgového kmeňa), ako aj so subkortikálnymi jadrami (pallidus, putamen, caudate nucleus) a s novou kôrou mozgových hemisfér, predovšetkým s kôrou spánkového a predného laloku.
Napriek fylogenetickým, morfologickým a cytoarchitektonickým rozdielom sú mnohé zo spomínaných štruktúr (limbická oblasť, centrálne a mediálne štruktúry talamu, hypotalamu, retikulárnej formácie mozgového kmeňa) zvyčajne zaradené do tzv. limbicko-retikulárny komplex, ktorý pôsobí ako zóna integrácie mnohých funkcií, zabezpečujúca organizáciu polymodálnych, holistických reakcií tela na rôzne vplyvy, čo je obzvlášť výrazné v stresových situáciách.
Štruktúry limbicko-retikulárneho komplexu majú veľké množstvo vstupy a výstupy, ktorými prechádzajú bludné kruhy početných aferentných a eferentných spojení, ktoré zabezpečujú kombinované fungovanie útvarov zahrnutých v tomto komplexe a ich interakcia so všetkými časťami mozgu, vrátane mozgovej kôry.
V štruktúrach limbicko-retikulárneho komplexu dochádza ku konvergencii senzitívnych impulzov, ktoré sa vyskytujú v intero- a exteroreceptoroch, vrátane receptorových polí zmyslových orgánov. Na tomto základe v limbicko-retikulárnom komplexe primárna syntéza informácií o stave vnútorného prostredia organizmu, ako aj o faktoroch pôsobiacich na organizmus vonkajšie prostredie a formujú sa elementárne potreby, biologické motivácie a sprievodné emócie.
Limbicko-retikulárny komplex určuje stav emocionálnej sféry, podieľa sa na regulácii vegetatívno-viscerálnych vzťahov zameraných na udržiavanie relatívnej stálosti vnútorného prostredia (homeostázy), ako aj zásobovania energiou a korelácie pohybových aktov. Od jeho stavu závisí úroveň vedomia, možnosť automatizovaných pohybov, činnosť motorických a mentálnych funkcií, reč, pozornosť, schopnosť orientácie, pamäť, zmena bdelosti a spánku.
Poškodenie štruktúr limbicko-retikulárneho komplexu môže byť sprevádzané rôznymi klinickými príznakmi: výrazné zmeny v emocionálnej sfére trvalého a paroxyzmálneho charakteru, anorexia alebo bulímia, sexuálne poruchy, poruchy pamäti, najmä príznaky Korsakoffovho syndrómu, pri ktorých pacient stráca schopnosť zapamätať si aktuálne udalosti (uchováva aktuálne udalosti v pamäti nie dlhšie ako 2 minúty), vegetatívno-endokrinné poruchy, poruchy spánku, psychosenzorické poruchy vo forme ilúzií a halucinácií, zmeny vedomia, klinické prejavy akinetického mutizmu, epileptické záchvaty.
K dnešnému dňu sa uskutočnilo veľké množstvo štúdií o štúdiu morfológie, anatomických vzťahov, funkcie limbickej oblasti a iných štruktúr zahrnutých do limbicko-retikulárneho komplexu, avšak fyziológia a znaky klinického obrazu jeho lézie dnes je ešte potrebné do značnej miery objasniť. Väčšina informácií o jeho funkcii, najmä funkcie parahipokampálnej oblasti, získané pri pokusoch na zvieratách metódy podráždenia, extirpácie alebo stereotaxie. Získané týmto spôsobom výsledky vyžadujú opatrnosť pri extrapolácii na ľudí. Zvláštny význam majú klinické pozorovania pacientov s léziami mediobazálnych oblastí mozgovej hemisféry.
V 50-60 rokoch XX storočia. pri rozvoji psychochirurgie sa objavili správy o liečbe pacientov s nevyliečiteľnou mentálne poruchy a syndróm chronickej bolesti bilaterálnou cingulotómiou (disekcia cingulárneho gyru), zatiaľ čo regresia úzkosti, obsedantných stavov, psychomotorickej agitácie, bolestivých syndrómov bola zvyčajne zaznamenaná, čo bolo uznané ako dôkaz zapojenia cingulárneho gyru do vytvárania emócií a bolesť. Zároveň bicingulotómia viedla k hlbokým poruchám osobnosti, k dezorientácii, zníženiu kritickosti vlastného stavu a eufórii.
Analýza 80 overených klinických prípadov hipokampálnych lézií na základe Neurochirurgického inštitútu Ruskej akadémie lekárskych vied je uvedená v monografii N.N. Bragina (1974). Autor prichádza k záveru, že temporálny mediobazálny syndróm zahŕňa viscerovegetatívne, motorické a duševné poruchy, prejavujúce sa zvyčajne komplexne. Všetka rozmanitosť klinických prejavov N.N. Bragin redukuje na dva hlavné multifaktoriálne varianty patológie s prevahou "dráždivých" a "inhibičných" javov.
Prvý z nich zahŕňa emocionálne poruchy sprevádzané motorickou úzkosťou (zvýšená excitabilita, upovídanost, nervozita, pocit vnútornej úzkosti), záchvaty strachu, vitálna úzkosť, rôzne viscerovegetatívne poruchy (zmeny pulzu, dýchania, gastrointestinálne poruchy, horúčka, zvýšené potenie atď.). U týchto pacientov sa na pozadí neustáleho motorického nepokoja často vyskytli záchvaty motorickej excitácie.
niya. EEG tejto skupiny pacientov sa vyznačovalo miernymi cerebrálnymi zmenami smerom k integrácii (zrýchlený a vypointovaný alfa rytmus, difúzne beta oscilácie). Opakované aferentné stimuly vyvolali jasné EEG odpovede, ktoré na rozdiel od normálnych nezmizli, keď boli stimuly opakovane prezentované.
Druhý („inhibičný“) variant mediobazálneho syndrómu je charakterizovaný emocionálnymi poruchami vo forme depresie s motorickou retardáciou (potlačená nálada, ochudobnenie a spomalenie tempa duševných procesov, zmeny motoriky, pripomínajúce akineticko-rigidný syndróm typu.Viscerovegetatívne paroxyzmy zaznamenané v prvej skupine sú menej charakteristické.EEG pacientov v tejto skupine bolo charakterizované cerebrálnymi zmenami, prejavujúcimi sa prevahou pomalých foriem aktivity (nepravidelný, pomalý alfa rytmus, skupiny kmitov theta, difúzne delta vlny ).Ostrý pokles EEG reaktivity pritiahol pozornosť.
Medzi týmito dvoma krajnými variantmi boli aj stredné s prechodnými a zmiešanými kombináciami jednotlivých symptómov. Niektoré z nich sa teda vyznačujú relatívne slabými známkami rozrušenej depresie so zvýšenou motorickou aktivitou a únavou, s prevahou senestopatických pocitov, podozrievavosťou, ktorá u niektorých pacientov dosahuje paranoidné stavy, a hypochondrickým delíriom. Druhá stredná skupina sa vyznačovala extrémnou intenzitou depresívnych symptómov na pozadí stuhnutosti pacienta.
Tieto údaje nám umožňujú hovoriť o duálnom (aktivujúcom a inhibičnom) vplyve hipokampu a iných štruktúr limbickej oblasti na behaviorálne reakcie, emócie, mentálny stav a bioelektrickú aktivitu kôry. V súčasnosti by sa komplexné klinické syndrómy tohto typu nemali považovať za primárne fokálne. Skôr by sa o nich malo uvažovať vo svetle predstáv o viacúrovňový systém organizácia mozgovej činnosti.
S.B. Buklina (1997) citoval údaje z prieskumu 41 pacientov s arteriovenóznymi malformáciami v oblasti gyrus cingulate. Pred operáciou malo 38 pacientov poruchy pamäti v popredí a päť z nich malo znaky Korsakoffovho syndrómu, u troch pacientov Korsakoffov syndróm vznikol po operácii, pričom závažnosť nárastu porúch pamäti korelovala so stupňom deštrukcie gyrus cingulate. samotnej, ako aj s postihnutím patologického procesu priľahlých štruktúr corpus callosum, pričom amnesický syndróm nezávisel od strany lokalizácie malformácie a jej lokalizácie po dĺžke gyrus cingulate.
Hlavnými charakteristikami zistených amnestických syndrómov boli poruchy reprodukcie sluchovo-rečových podnetov, porušenie selektivity stôp vo forme inklúzií a kontaminácií a neschopnosť zachovať význam pri prenose príbehu. U väčšiny pacientov bola kritickosť hodnotenia ich stavu znížená. Autor zaznamenal podobnosť týchto porúch s amnestickými defektmi u pacientov s frontálnymi léziami, čo možno vysvetliť prítomnosťou spojení medzi gyrus cingulate a frontálnym lalokom.
Viac rozsiahle patologické procesy v limbickej oblasti spôsobujú výrazné poruchy vegetatívno-viscerálnych funkcií.
corpus callosum(corpus callosum)- najväčšia komisura medzi mozgovými hemisférami. Jeho predné časti, najmä koleno corpus callosum
telo (genu corporis callosi), spájajú predné laloky, stredné časti - kmeň corpus callosum (truncus corporis callosi)- zabezpečujú komunikáciu medzi temporálnymi a parietálnymi časťami hemisfér, zadnými časťami, najmä hrebeňom corpus callosum (splenium corporis callosi), spojte okcipitálne laloky.
Lézie corpus callosum sú zvyčajne sprevádzané poruchami duševného stavu pacienta. Zničenie jeho prednej časti vedie k rozvoju „frontálnej psychiky“ (spontánnosť, porušovanie akčného plánu, správanie, kritika, charakteristika frontálny kalous syndróm - akinéza, amimia, spontánnosť, astázia-abázia, apraxia, úchopové reflexy, demencia). Prerušenie spojení medzi parietálnymi lalokmi vedie k perverzii pochopenie "telesné plány" a vzhľad apraxie väčšinou v ľavej ruke. Disociácia temporálnych lalokov sa môže prejaviť narušenie vnímania vonkajšieho prostredia, strata správnej orientácie v ňom (amnestické poruchy, konfabulácie, syndróm už videného atď.). Patologické ložiská v zadných častiach corpus callosum sú často charakterizované príznakmi vizuálnej agnózie.
14.6. ARCHITEKTONIKA MOZGOVEJ KÔRY
Štruktúra mozgovej kôry je heterogénna. Menej zložité v štruktúre, skoro vznikajúce v procese fylogenézy starodávna kôra (archiokortex) a stará kôra (paleokortex), súvisiace väčšinou do limbického laloku mozog. Väčšia časť mozgovej kôry (95,6 %) je v dôsledku neskoršej fylogenetickej tvorby tzv. nová kôra (neokortex) a má oveľa zložitejšiu viacvrstvovú štruktúru, ale aj heterogénnu vo svojich rôznych zónach.
Vzhľadom k tomu, že architektonika kôry je v určitom spojení s jej funkciou, jeho štúdiu bola venovaná veľká pozornosť. Jedným zo zakladateľov doktríny cytoarchitektoniky kôry bol V.A. Betz (1834-1894), ktorý v roku 1874 prvýkrát opísal veľké pyramídové bunky motorickej kôry (Betzove bunky) a určil princípy delenia mozgovej kôry na hlavné oblasti. V budúcnosti veľký prínos k rozvoju teórie štruktúry kôry urobili mnohí výskumníci - A. Campbell (A. Cambell), E. Smith (E. Smith), K. Brodmann (K. Brodmann ), Oscar Vogt a Cecilia Vogt (O. Vogt, S. Vogt). Veľkú zásluhu na štúdiu architektoniky kôry majú pracovníci Ústavu mozgu Akadémie lekárskych vied (S.A. Sarkisov, N.I. Filimonov, E.P. Kononova atď.).
Hlavný typ štruktúry novej kôry (obr. 14.5), s ktorou sa porovnávajú všetky jej úseky – kôra pozostávajúca zo 6 vrstiev (homotypická kôra podľa Brodmana).
Vrstva I - molekulárna alebo zonálna, najpovrchnejšia, chudobná na bunky, jej vlákna majú smer, hlavne rovnobežný s povrchom kôry.
II vrstva - vonkajšia zrnitá. Pozostáva z veľkého počtu husto usporiadaných malých granulovaných nervových buniek.
III vrstva - malé a stredné pyramídy, najširšie. Pozostáva z pyramídových buniek, ktorých veľkosti nie sú rovnaké, čo umožňuje rozdeliť túto vrstvu na podvrstvy vo väčšine kortikálnych polí.
IV vrstva - vnútorná granulovaná. Skladá sa z husto usporiadaných malých buniek - zŕn okrúhleho a hranatého tvaru. Táto vrstva je najvariabilnejšia
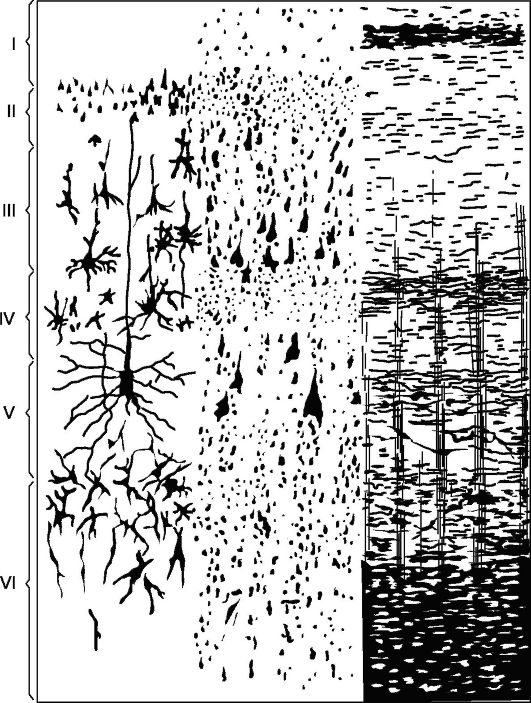
Ryža. 14.5.Cytoarchitektonika a myeloarchitektonika motorickej zóny mozgovej kôry.
Vľavo: I - molekulárna vrstva; II - vonkajšia zrnitá vrstva; III - vrstva malých a stredných pyramíd; IV - vnútorná zrnitá vrstva; V - vrstva veľkých pyramíd; VI - vrstva polymorfných buniek; vpravo - prvky myeloarchitektoniky.
v niektorých poliach (napr. pole 17) je rozdelená na podvrstvy, na niektorých miestach sa prudko stenčuje a dokonca úplne zaniká.
V vrstva - veľké pyramídy alebo gangliové. Obsahuje veľké pyramídové bunky. V niektorých oblastiach mozgu je vrstva rozdelená na podvrstvy, v motorickej zóne sa skladá z troch podvrstiev, z ktorých stred obsahuje Betzove obrie pyramídové bunky, dosahujúce v priemere 120 mikrónov.
VI vrstva - polymorfné bunky alebo multiformné. Pozostáva prevažne z trojuholníkových vretenovitých buniek.
Štruktúra mozgovej kôry má veľké množstvo variácií v dôsledku zmien hrúbky jednotlivých vrstiev, stenčovania alebo vymiznutia, resp.
naopak, zhrubnutie a rozdelenie na podvrstvy niektorých z nich (heterotypické zóny podľa Brodmana).
Kôra každej mozgovej hemisféry je rozdelená do niekoľkých oblastí: okcipitálna, horná a dolná parietálna, postcentrálna, centrálna gyri, precentrálna, frontálna, temporálna, limbická, ostrovná. Každý z nich v súlade s charakteristikami rozdelené do niekoľkých polí, navyše každé pole má svoje konvenčné poradové označenie (obr. 14.6).
Štúdium architektoniky mozgovej kôry spolu s fyziologickými, vrátane elektrofyziologických štúdií a klinických pozorovaní výrazne prispelo k riešeniu problému rozloženia funkcií v kôre.
14.7. PROJEKČNÉ A ASOCIAČNÉ POLIA KORTUSU
V procese rozvíjania doktríny o úlohe mozgovej kôry a jej jednotlivých úsekov pri výkone určitých funkcií existovali rôzne, niekedy aj opačné pohľady. Panoval teda názor o prísne lokálnom zastúpení všetkých ľudských schopností a funkcií v mozgovej kôre až po najzložitejšie, mentálne (lokalizácia, psychomorfologizmus). Oponoval mu iný názor o absolútnej funkčnej rovnocennosti všetkých častí mozgovej kôry. (ekvipotencializmus).
Významný príspevok k teórii lokalizácie funkcií v mozgovej kôre priniesol I.P. Pavlov (1848-1936). Vybral projekčné zóny kôry (kortikálne konce analyzátorov určité typy citlivosť) a asociatívne zóny umiestnené medzi nimi, študovali procesy inhibície a excitácie v mozgu, ich vplyv na funkčný stav mozgovej kôry. Rozdelenie kôry na projekčné a asociatívne zóny prispieva k pochopeniu organizácie práce mozgovej kôry a je opodstatnené pri riešení praktických problémov, najmä v aktuálnej diagnostike.
projekčné zóny poskytujú predovšetkým jednoduché špecifické fyziologické úkony, predovšetkým vnímanie vnemov určitej modality. Projekčné dráhy, ktoré sa k nim približujú, spájajú tieto zóny s receptorovými územiami na periférii, ktoré sú s nimi vo funkčnej korešpondencii. Príklady projekčných kortikálnych zón sú oblasť zadného centrálneho gyrusu už opísaná v predchádzajúcich kapitolách (zóna všeobecné typy citlivosť) alebo oblasť ostrohy umiestnenej na mediálnej strane okcipitálneho laloku (projekčná vizuálna zóna).
Asociačné zóny kôra nemá priame spojenie s perifériou. Nachádzajú sa medzi projekčnými zónami a majú početné asociatívne väzby s týmito projekčnými zónami as inými asociačnými zónami. Funkciou asociatívnych zón je vykonávať vyššiu analýzu a syntézu mnohých elementárnych a zložitejších komponentov. Tu v podstate dochádza k pochopeniu informácií vstupujúcich do mozgu, k formovaniu myšlienok a konceptov.
G.I. Polyakov v roku 1969 na základe porovnania architektoniky ľudskej mozgovej kôry a niektorých zvierat zistil, že asociatívne
Ryža. 14.6.Architektonické polia mozgovej kôry [podľa Brodmana]. a - vonkajší povrch; b - mediálna plocha.
zóny v mozgovej kôre človeka sú 50 %, v kôre vyšších (humanoidných) opíc - 20 %, u nižších opíc je toto číslo 10 % (obr. 14.7). Medzi asociačné oblasti kôry ľudského mozgu, ten istý autor navrhol izolovať sekundárne a terciárne odbory. Sekundárne asociatívne polia susedia s projekčnými. Vykonávajú analýzu a syntézu elementárnych vnemov, ktoré si stále zachovávajú špecifickú orientáciu.
Terciárne asociatívne polia sa nachádzajú prevažne medzi sekundárnymi a sú prekrývajúcimi sa zónami susedných území. Súvisia predovšetkým s analytickou činnosťou kôry, ktorá zabezpečuje najvyššie mentálne funkcie človeka v jeho najzložitejších intelektuálnych a rečových prejavoch. Funkčná zrelosť terciárneho as-
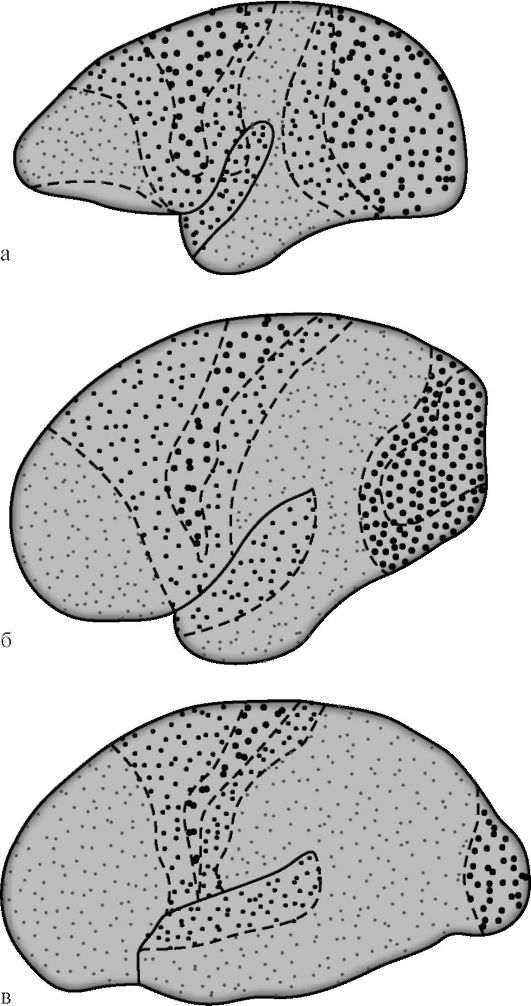
Ryža. 14.7. Diferenciácia projekčných a asociačných oblastí mozgovej kôry počas evolúcie primátov [podľa G.I. Polyakov]. a - mozog nižšej opice; b - mozog vyššej opice; c - ľudský mozog. Veľké bodky označujú projekčné zóny, malé bodky - asociatívne. U nižších opíc zaberajú asociatívne zóny 10% plochy kôry, u vyšších - 20%, u ľudí - 50%.
sociálne polia mozgovej kôry sa vyskytuje najneskoršie a to len v priaznivom sociálnom prostredí. Na rozdiel od iných kortikálnych polí sa terciárne polia pravej a ľavej hemisféry vyznačujú výrazným funkčná asymetria.
14.8. TOPICKÁ DIAGNOSTIKA LEZIÍ MOZGOVEJ KÔRY
14.8.1. Prejavy poškodenia projekčných zón mozgovej kôry
V kôre každej mozgovej hemisféry za centrálnym gyrusom je 6 projekčných zón.
1. V prednej časti parietálneho laloku, v oblasti zadného centrálneho gyru (cytoarchitektonické polia 1, 2, 3) Nachádza projekčná zóna všeobecných typov citlivosti(obr. 14.4). Oblasti kôry, ktoré sa tu nachádzajú, dostávajú citlivé impulzy prichádzajúce pozdĺž projekčných dráh všeobecných typov citlivosti z receptorového aparátu opačnej polovice tela. Čím vyššia je plocha tejto projekčnej zóny kôry, tým nižšie sú umiestnené časti opačnej polovice tela, ktoré má projekčné spojenia. Časti tela s rozsiahlym príjmom (jazyk, palmárna plocha ruky) zodpovedajú neadekvátne veľkým častiam plochy projekčných zón, zatiaľ čo iné časti tela (proximálne končatiny, trup) majú malú plochu kortikálnej zastupovanie.
Podráždenie patologickým procesom kortikálnej zóny všeobecných typov citlivosti vedie k záchvatu parestézie v častiach tela zodpovedajúcich podráždeným oblastiam mozgovej kôry (citlivý Jacksonov záchvat), ktorý sa môže zmeniť na sekundárny generalizovaný paroxyzmus. Porážka kortikálneho konca analyzátora všeobecných typov citlivosti môže spôsobiť rozvoj hypalgézie alebo anestézie v zodpovedajúcej zóne opačnej polovice tela, zatiaľ čo miesto hypestézie alebo anestézie môže byť vertikálne obehové alebo radikulárne. segmentový typ. V prvom prípade sa porucha citlivosti prejavuje na opačnej strane k patologickému ložisku v oblasti pier, palca alebo v distálnej časti končatiny s kruhovým okrajom, niekedy ako ponožka alebo rukavica. V druhom prípade má zóna narušenia citlivosti tvar pásika a nachádza sa pozdĺž vnútorného alebo vonkajšieho okraja ramena alebo nohy; vysvetľuje to skutočnosť, že vnútorná strana končatín je prezentovaná v prednej časti a vonkajšia strana - v zadných častiach projekčnej zóny analyzátora všeobecných typov citlivosti.
2. Zóna vizuálnej projekcie Nachádza v kortexe mediálneho povrchu okcipitálneho laloku v oblasti ostrohy (pole 17). V tomto poli dochádza k stratifikácii IV (vnútornej zrnitej) vrstvy kortexu so zväzkom myelínových vlákien do dvoch podvrstiev. Oddelené úseky poľa 17 prijímajú impulzy z určitých úsekov homonymných polovíc sietníc oboch očí; pričom impulzy prichádzajúce zo spodných častí homonymných polovíc sietníc dosahujú
kôra spodnej pery ostrohovej drážky a impulzy prichádzajúce z horných častí sietníc smerujú do kôry jej hornej pery.
Porážka patologického procesu zóny vizuálnej projekcie vedie k tomu, že sa na opačnej strane objaví kvadrant alebo úplná homonymná hemianopia na strane opačnej k patologickému zameraniu. Obojstranné poškodenie kortikálnych polí 17 alebo k nim vedúcich projekčných zrakových dráh môže viesť k úplnej slepote. Podráždenie kôry zóny vizuálnej projekcie môže spôsobiť výskyt vizuálnych halucinácií vo forme fotopsií v zodpovedajúcich častiach opačných polovíc zorných polí.
3. Oblasť projekcie sluchu Nachádza v kôre Heschlových zákrut na spodnom okraji bočnej (Sylvian) brázdy (polia 41 a 42), ktoré sú v skutočnosti súčasťou horného temporálneho gyru. Podráždenie tejto zóny kôry môže spôsobiť výskyt sluchových halucinácií (útoky pocitového hluku, zvonenie, pískanie, bzučanie atď.). Zničenie zóny sluchovej projekcie na jednej strane môže spôsobiť určitú stratu sluchu v oboch ušiach, vo väčšej miere naopak vzhľadom na patologické zameranie.
4 a 5. Čuchové a chuťové projekčné zóny sú na mediálnom povrchu klenutého gyrusu (limbickej oblasti) mozgu. Prvý z nich sa nachádza v parahipokampálnom gyre (pole 28). Projekčná zóna chuti je zvyčajne lokalizovaná v kortexe operkulárnej oblasti (pole 43). Podráždenie projekčných zón vône a chuti môže spôsobiť ich zvrátenie alebo viesť k rozvoju zodpovedajúcich čuchových a chuťových halucinácií. Jednostranná strata funkcie projekčných zón vône a chuti môže spôsobiť mierny pokles vône a chuti na oboch stranách. Obojstranná deštrukcia kortikálnych koncov tých istých analyzátorov sa prejavuje absenciou vône a chuti na oboch stranách, resp.
6. Vestibulárna projekčná zóna. Jeho lokalizácia nie je špecifikovaná. Zároveň je známe, že vestibulárny aparát má početné anatomické a funkčné spojenia. Je možné, že lokalizácia zastúpenia vestibulárneho systému v kortexe ešte nie je objasnená, pretože je polyfokálny. N.S. Blagoveshchenskaya (1981) verí, že v mozgovej kôre sú vestibulárne projekčné zóny reprezentované niekoľkými anatomickými a funkčnými interakčnými komplexmi, ktoré sa nachádzajú v poli 8, na križovatke frontálnych, temporálnych a parietálnych lalokov a v zóne centrálneho gyru. , pričom sa predpokladá, že každá z týchto oblastí kôry plní svoje vlastné funkcie. Pole 8 je ľubovoľným centrom pohľadu, jeho podráždenie spôsobuje otočenie pohľadu opačným smerom ako je patologické ohnisko, zmeny v rytme a povahe experimentálneho nystagmu, najmä krátko po epileptickom záchvate. V kôre spánkového laloka sa nachádzajú útvary, ktorých podráždenie spôsobuje závraty, ktoré sa prejavujú najmä pri epilepsii spánkového laloku; porážka oblastí reprezentácie vestibulárnych štruktúr v kôre centrálneho gyri ovplyvňuje stav tonusu priečne pruhovaných svalov. Klinické pozorovania naznačujú, že jadrovo-kortikálne vestibulárne dráhy spôsobujú čiastočnú dekusáciu.
Je potrebné zdôrazniť, že príznaky podráždenia uvedených projekčných zón môžu byť prejavom aury epileptického záchvatu zodpovedajúceho charakteru.
I.P. Pavlov považoval za možné uvažovať o kôre precentrálneho gyru, ktorá ovplyvňuje motorické funkcie a svalový tonus prevažne opačnej polovice tela, s ktorou je spojená predovšetkým kortikálno-nukleárnymi a kortikálno-spinálnymi (pyramídovými) dráhami, ako projekčná zóna tzv motorový analyzátor. Táto zóna zaberá v prvom rade pole 4, na ktoré sa v obrátenej podobe premieta opačná polovica tela. Toto pole obsahuje väčšinu obrovských pyramídových buniek (Betzove bunky), ktorých axóny tvoria 2-2,5 % všetkých vlákien pyramídovej dráhy, ako aj stredné a malé pyramídové bunky, ktoré spolu s axónmi toho istého bunky nachádzajúce sa v susednom poli 4 rozsiahlejšie pole 6, sa podieľajú na realizácii monosynaptických a polysynaptických kortikálno-svalových spojení. Monosynaptické spojenia zabezpečujú predovšetkým rýchle a presné cielené akcie v závislosti od kontrakcií jednotlivých priečne pruhovaných svalov.
Poškodenie spodných častí motorickej zóny zvyčajne vedie k vývoju na opačnej strane brachiofaciálny (plecia na tvár) syndróm alebo linguofaciobrachiálny syndróm, ktoré sú často pozorované u pacientov s poruchou cerebrálnej cirkulácie v povodí strednej mozgovej tepny, zatiaľ čo kombinovaná paréza svalov tváre, jazyka a paže, najmä ramena v centrálnom type.
Podráždenie kôry motorickej zóny (polia 4 a 6) vedie k vzniku kŕčov vo svaloch premietnutých do tejto zóny resp. svalové skupiny. Častejšie ide o lokálne kŕče typu Jacksonovej epilepsie, ktoré sa môžu pretransformovať do sekundárneho generalizovaného epileptického záchvatu.
14.8.2. Prejavy poškodenia asociačných polí mozgovej kôry
Medzi projekčnými zónami kôry sú asociačné polia. Dostávajú impulzy najmä z buniek projekčných zón kôry. V asociatívnych poliach prebieha analýza a syntéza informácií, ktoré prešli primárnym spracovaním v projekčných poliach. Asociatívne zóny kôry horného parietálneho laloku poskytujú syntézu elementárnych pocitov, v súvislosti s tým také zložité typy citlivosti, ako je zmysel pre lokalizáciu, pocit hmotnosti, dvojrozmerný priestorový zmysel, ako aj komplexné tu sa tvoria kinestetické vnemy.
V oblasti interparietálneho sulcus sa nachádza asociatívna zóna, ktorá zabezpečuje syntézu vnemov vychádzajúcich z častí vlastného tela. Poškodenie tejto oblasti kôry vedie k autopagnóza, tie. k nerozpoznaniu alebo ignorovaniu častí vlastného tela, alebo k pseudomelia pocit, že máte ďalšiu ruku alebo nohu, a anozognózia - nedostatočná informovanosť o telesnom defekte, ktorý vznikol v súvislosti s ochorením (napríklad ochrnutie alebo paréza končatiny). Zvyčajne sa všetky typy autopagnózie a anozognózie vyskytujú, keď je patologický proces umiestnený vpravo.
Porážka dolného parietálneho lalôčika sa môže prejaviť poruchou syntézy elementárnych vnemov alebo neschopnosťou porovnať syntetizované komplexné vnemy s „kedysi bolo vo vnímaní podobných
rovnakým spôsobom, na základe výsledkov ktorých dochádza k uznaniu “(V.M. Bekhterev). To sa prejavuje porušením dvojrozmerného priestorového zmyslu (grafestézia) a trojrozmerného priestorového zmyslu (stereognóza) - astereognózy.
Pri poškodení premotorických zón frontálneho laloka (polia 6, 8, 44) väčšinou dochádza k frontálnej ataxii, pri ktorej je narušená syntéza aferentných impulzov (kinestetická aferentácia), signalizujúcich polohu častí tela v priestore, ktorá zmeny počas vykonávaných pohybov.
V prípade narušenia funkcie kôry predných častí čelového laloka, ktorá má spojenia s opačnou hemisférou mozočka (fronto-mostíkové-cerebelárne spojenia), dochádza k poruchám statokinetiky na opačnej strane patologického ložiska. (frontálna ataxia). Zvlášť zreteľné sú porušenia neskoro sa rozvíjajúcich foriem statokinetiky - vzpriamené státie a vzpriamená chôdza. V dôsledku toho má pacient neistotu, neistotu chôdze. Pri chôdzi sa jeho telo nakláňa dozadu. (Hennerov znak) dáva nohy do priamej línie (líška chôdza) niekedy pri chôdzi dochádza k "opleteniu" nôh. U niektorých pacientov s poškodením predných častí predných lalokov sa vyvinie zvláštny jav: pri absencii paralýzy a parézy a zachovanej schopnosti pohybovať nohami v plnom rozsahu pacienti nemôžu stáť (astasia) a chodiť (abázia).
Porážka asociačných zón kôry je často charakterizovaná vývojom klinických prejavov porušenia vyšších mentálnych funkcií (pozri kapitolu 15).








