1. Przypomnij sobie z kursu zoologii, które części mózgu mają wszystkie kręgowce. Z jakiego działu pochodzą półkule mózgowe? W jakiej grupie zwierząt pojawiają się po raz pierwszy?
Wszystkie kręgowce charakteryzują się obecnością 5 części mózgu: rdzeń tyłomózgowie, śródmózgowie, międzymózgowie, przodomózgowie.
Ogólny przegląd budowy półkul mózgowych
Niezrozumienie mózgu konia Nierzadko słyszy się następujące stwierdzenia: W jednym koniu są dwa konie: prawy koń i niezawodny lewy koń oraz nieprzewidywalny prawy koń, odnosząc się w ten sposób do dwóch półkul mózgu: lewej półkuli dla logika. Nigdy nie powinieneś jeździć konno z prawą półkulą i najpierw uczyć konia używania lewej półkuli. Koń ma dwie świadomości wzrokowe. Koń nie myśli, reaguje instynktownie. Ale te twierdzenia nie są oparte na dowodach naukowych.
Półkule mózgowe są pochodnymi przodomózgowia i po raz pierwszy pojawiają się u płazów, ale są słabo rozwinięte, kora mózgowa jest praktycznie nieobecna.
2. Opisz strukturę półkule osoba. Narysuj schematyczny przekrój półkul mózgowych, wskazując na rysunku istotę szarą kory, istotę szarą jąder, istotę białą i komory.
Spojrzenie na historię ewolucji ssaków w hominidy i miejsce konia w tej historii jest sposobem na argumentację przeciwko tym błędom w analizie zachowania koni. Klasa ssaków ma kilka podziałów lub rzędów, takich jak gryzonie, zwierzęta mięsożerne, owadożerne, kopytne i naczelne.
Ssaki to zwierzęta charakteryzujące się produkcją mleka z gruczołów sutkowych samicy. Koń należy do klasy ssaków i do rzędu kopytnych, ponieważ brzegi palców pokryte są kopytem, a kończyny opierają się na ostatniej falangi. Koń został jednak udomowiony dopiero później, pierwsze ślady udomowienia datują się na okres od 500 do tysiąca lat. Ewolucja mózgu u niektórych dorosłych ssaków. całkowita masa mózgu w gramach.
U ludzi przodomózgowie są reprezentowane przez dwie półkule i ciało modzelowate łączące półkule. Duże półkule: prawa i lewa - pokrywają środkową i międzymózgowia i stanowią do 80% masy mózgu osoby dorosłej. Na powierzchni każdej półkuli znajduje się wiele bruzd i zwojów. Istnieją 4 główne bruzdy (centralna, boczna i ciemieniowo-potyliczna), które dzielą każdą półkulę na płaty. Warstwa powierzchniowa składa się z istoty szarej (kory), poniżej znajduje się istota biała, składająca się z aksonów komórek nerwowych, których ciała leżą w korze lub przenoszą informacje do komórek kory. Duże skupiska istoty szarej (jądra podkorowe) i jamy (komory boczne) leżą w grubości istoty białej. Przez komory mózgu i kanał centralny rdzeń kręgowy krąży płyn mózgowo-rdzeniowy, który zapewnia odżywienie struktur podkorowych.
Całkowita masa mózgu wyrażona jako procent masy ciała. Naczelne i hominidy. W obrębie klasy ssaków istnieje również rząd naczelnych, który pojawia się na początku trzeciorzędu. Wśród tych naczelnych są pongidae lub małpy człekokształtne i hominidy. Hominid to rodzina dwunożnych naczelnych, czyli austratopitów, takich jak Lucy, a później samce.
Ewolucja mózgu Badanie ewolucji organizmów żywych na przestrzeni milionów lat od pierwszych ssaków do ludzi umożliwiło obserwację główny trend mózgu do zwiększenia objętości na całym świecie i na korzyść określonych obszarów mózgu. U ssaków ten wzrost był głównie na korzyść półkul mózgowych i móżdżku, tak że w przeciwieństwie do ptaków i gadów, tylko ssaki rozwinęły neorektorię z 6 warstwami komórek, ta nocort stanowi ponad połowę objętości mózgu u najbardziej zaawansowanych ssaków.
3. Czym jest kora mózgowa? Gdzie to jest?
Warstwa powierzchniowa istoty szarej półkul mózgowych nazywana jest korą. Kora składa się z kilku warstw ciał neuronów, różniących się budową i funkcją. Uważa się, że w jego składzie znajduje się około 12-18 miliardów komórek, grubość 1,5-4,5 mm, a powierzchnia 1,7-2,5 tys. cm2.
Struktura płata czołowego
W ewolucji hominidów zaobserwowano również wzrost pojemności czaszki poprzez stopniowy wzrost mózgu i móżdżku. Jedna z funkcji ludzki mózg w przeciwieństwie do innych ssaków jest jego asymetria anatomiczna i funkcjonalna. Na przykład duża część lewej półkuli specjalizuje się w wytwarzaniu i rozumieniu języka, pisaniu i czytaniu. Prawa półkula jest bardziej konkretnie związana z orientacją przestrzenną w wykrywaniu formy wizualne i wiedza muzyczna. Lewa półkula jest bardziej dominująca dla procesów, logicznych, analitycznych i prawa dla procesów holistycznych.
4. Wyjaśnij znaczenie bruzd i zwojów na powierzchni półkul mózgowych.
Bruzdy i zakręt znacznie zwiększają powierzchnię kory mózgowej, według niektórych źródeł nawet 10-12 razy.
5. Jaka jest funkcja istoty białej półkul mózgowych?
Istota biała tworzy ścieżki łączące korę i korę z resztą układu nerwowego.
Zatem asymetria mózgu jest funkcja ludzki, a także brak niezwykłej sieci, u małp, wciąż mających symetryczne mózgi, starożytny norcort zachowujący swoje symetryczne funkcjonowanie, więc logika nie powinna być implikowana. u konia jedna z dwóch półkul pełni pewną funkcję, mówią niektórzy z nowych mistrzów.
Ale nawet u ludzi półkula mózgowa nigdy nie jest jedyną, która zakłóca tę funkcję, ale tylko wydaje się, że jest bardziej połączona, uwikłana w tę funkcję. W rzeczywistości podstawowe funkcje mózgu pozostają w dużej mierze rozłożone na wszystkie dwie półkule mózgu. W związku z tym ważne jest również, aby przypomnieć obecność struktury ciała modzelowatego zarówno u ludzi, jak iu koni. Ta struktura jest prawdziwe połączenie między dwiema komunikującymi się ze sobą półkulami. Powiedzieć zatem, że połączenie między półkulami mózgu konia jest całkowicie błędne.
6. Jakie płaty są izolowane w półkulach mózgowych?
Na półkulach rozróżnia się płaty czołowe, 2 ciemieniowe, 2 skroniowe i potyliczne
7. Rozróżnij pojęcia „płatów półkul mózgowych” i „stref półkul mózgowych”. Podaj przykłady, kiedy pasują, nie pasują.
Płaty półkul mózgowych są podziałem powierzchni kory zgodnie z zasadą anatomiczną: w każdej półkuli rozróżnia się płaty czołowe, potyliczne, ciemieniowe i skroniowe.
Strefy korowe - odcinek kory mózgowej, charakteryzujący się jednolitością struktury i pełnionych funkcji.
W naszym mózgu płaty i strefy nie pokrywają się, jeden płat składa się z kilku stref. Płat potyliczny składa się z obszarów widzenia i rozpoznawania wizualnego. W płacie skroniowym i obok niego znajdują się strefy węchu, słuchu i smaku.
8. Podczas badania niewidomego pacjenta stwierdzono, że jego oczy i nerwy wzrokowe nie są uszkodzone. Dlaczego on nie widzi?
Niemożność widzenia może być spowodowana, oprócz uszkodzenia oczu lub nerwów wzrokowych, oraz uszkodzenia stref wzrokowych płatów potylicznych półkul mózgowych.
9. Korzystając z dodatkowych źródeł informacji, dowiedz się, czy funkcje lewej i prawej półkuli mózgu różnią się.
Istnieje „funkcjonalna asymetria” między prawą i lewą półkulą mózgu, to znaczy ich funkcje są różne. Zostało to udowodnione podczas eksperymentów z przecinaniem linii komunikacyjnych między półkulami (później takie operacje zaczęto przeprowadzać ze względów medycznych w niektórych schorzeniach, np. w chorobie Parkinsona). Osoby praworęczne mają lewą półkulę, podczas gdy osoby leworęczne mają prawą półkulę. Za to odpowiada prawa półkula kreatywne myslenie, stanowi podstawę kreatywności, podejmowania niestandardowych decyzji; Za rozpoznawanie twarzy odpowiada obszar wizualny prawej półkuli. Lewa półkula zapewnia logiczne rozumowanie i myślenie abstrakcyjne, zawiera ośrodki mowy ustnej i pisanej, formułowanie decyzji; strefa wizualna odpowiada za rozpoznawanie liter i cyfr. Dlatego żartobliwie praworęczni nazywani są matematykami, a leworęczni są artystami.
10. Istnieją codzienne koncepcje „męskiej logiki” i „kobiecej logiki”. Czy istnieją podstawy do takich różnic?
W strukturze zewnętrznej i wewnętrznej nie ma różnicy między mózgami mężczyzn i kobiet, ale pomimo tego, że struktura naszych mózgów jest prawie taka sama, jesteśmy indywidualni. Główne różnice w postrzeganiu świata według typu „męskiego” lub „żeńskiego” ujęte są w: wczesne dzieciństwo w zależności od naszego wychowania i pod wpływem hormonów gonad w okresie dorastania, ale wszystkie mają wyłącznie charakter psychologiczny.
14.1. POSTANOWIENIA OGÓLNE
Koniec mózgu (telencefalon) lub duży mózg (mózg), znajduje się w przestrzeni nadnamiotowej jamy czaszki składa się z dwóch dużych
półkule (gemispherium cerebralis),oddzielone głęboką podłużną szczeliną (fissura podłużnis cerebri), w którym zanurzony jest półksiężyc mózgu (fals cerebri) reprezentujący duplikację opony twardej. Duże półkule mózgu stanowią 78% jego masy. Każda z półkul mózgowych ma płaty: czołowy, ciemieniowy, skroniowy, potyliczny i limbiczny. Obejmują struktury międzymózgowia oraz pnia mózgu i móżdżku znajdujące się poniżej płaszcza móżdżku (podnamiotowo).
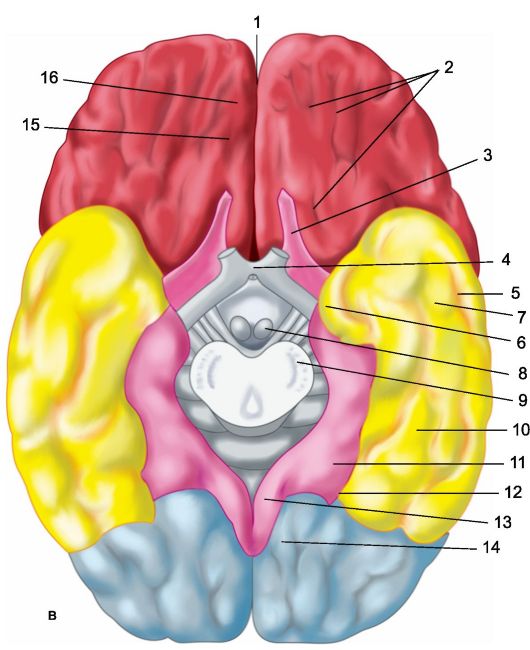
Każda z półkul mózgowych ma trzy powierzchnie: górny boczny lub wypukły (ryc. 14.1a), - wypukły, skierowany w stronę kości sklepienia czaszki; wewnętrzne (ryc. 14.1b), przylegające do dużego wyrostka sierpowatego i dolne lub podstawne (ryc. 14.1c), powtarzające relief podstawy czaszki (wgłębienia przednie i środkowe) oraz wypust móżdżku. Na każdej półkuli wyróżnia się trzy krawędzie: górną, dolną wewnętrzną i dolną zewnętrzną oraz trzy bieguny: przedni (przedni), tylny (potyliczny) i boczny (skroniowy).
Wnęka każdej półkuli mózgowej jest boczna komora mózgu podczas gdy lewa komora boczna jest rozpoznawana jako pierwsza, prawa - druga. Komora boczna ma centralną część umieszczoną głęboko w płacie ciemieniowym (lobus ciemieniowy) i wystają z niego trzy rogi: róg przedni penetruje płat czołowy (lobus czołowy), niższy - do doczesnego (lobus temporalis), tylny - w potylicy (lobus potyliczny). Każda z komór bocznych komunikuje się z trzecią komorą mózgu przez komorę międzykomorową otwór Monroe.
Centralne odcinki przyśrodkowej powierzchni obu półkul są połączone spoidłami mózgowymi, z których najbardziej masywnym jest ciało modzelowate i struktury międzymózgowia.
Kresomózgowia, podobnie jak inne części mózgu, składa się z istoty szarej i białej. Szara materia znajduje się w głębinach każdej półkuli, tworząc tam węzły podkorowe i wzdłuż obrzeży wolnych powierzchni półkuli, gdzie tworzy korę mózgową.
Główne zagadnienia związane ze strukturą, funkcjami jąder podstawnych oraz wariantami obrazu klinicznego w przypadku ich zajęcia omówiono w rozdziałach 5, 6. Kora mózgowa ma około
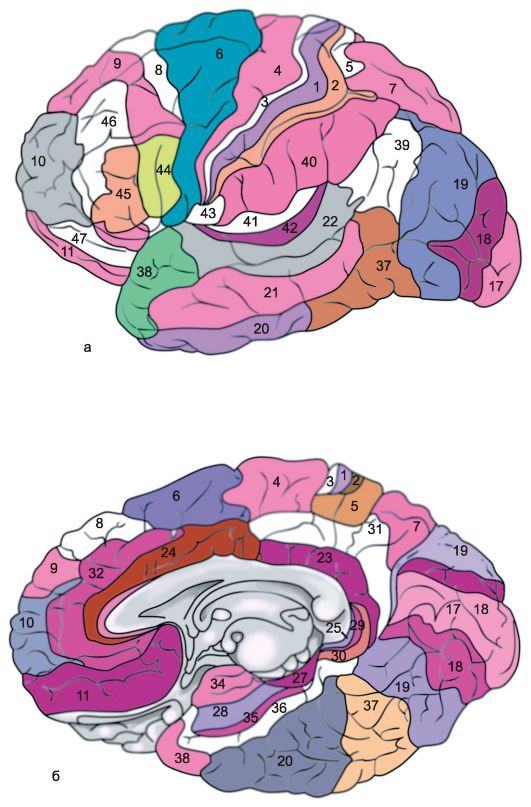
Ryż. 14.1.Półkule mózgu.
a - górna powierzchnia boczna lewej półkuli: 1 - bruzda środkowa; 2 - orbitalna część dolnego zakrętu czołowego; I - płat czołowy; 3 - zakręt przedśrodkowy; 4 - przedcentralna bruzda; 5 - lepszy zakręt czołowy; 6 - środkowy zakręt czołowy; 7 - nakrywkowa część dolnego zakrętu czołowego; 8 - dolny zakręt czołowy; 9 - boczna bruzda; II - płat ciemieniowy: 10 - zakręt postcentralny; 11 - bruzda postcentralna; 12 - rowek śródciemieniowy; 13 - zakręt nadbrzeżny; 14 - zakręt kątowy; III - płat skroniowy: 15 - górny zakręt skroniowy; 16 - górna bruzda skroniowa; 17 - środkowy zakręt skroniowy; 18 - środkowa bruzda skroniowa; 19 - dolny zakręt skroniowy; IV - płat potyliczny: b - przyśrodkowa powierzchnia prawej półkuli: 1 - płat przyśrodkowy, 2 - przedklinek; 3 - bruzda ciemieniowo-potyliczna; 4 - klin, 5 - zakręt językowy; 6 - boczny zakręt potyliczno-skroniowy; 7 - zakręt przyhipokampowy; 8 - hak; 9 - sklepienie; 10 - ciało modzelowate; 11 - lepszy zakręt czołowy; 12 - zakręt obręczy; c - dolna powierzchnia półkul mózgowych: 1 - podłużna szczelina międzypółkulowa; 2 - bruzdy orbitalne; 3 - nerw węchowy; 4 - skrzyżowanie optyczne; 5 - środkowa bruzda skroniowa; 6 - hak; 7 - dolny zakręt skroniowy; 8 - ciało wyrostka sutkowatego; 9 - podstawa pnia mózgu; 10 - boczny zakręt potyliczno-skroniowy; 11 - zakręt przyhipokampowy; 12 - rowek zabezpieczający; 13 - zakręt obręczy; 14 - zakręt językowy; 15 - rowek węchowy; 16 - bezpośredni zakręt.
3-krotność powierzchni półkul widocznych podczas badania zewnętrznego. Wynika to z faktu, że powierzchnia półkul mózgowych jest pofałdowana, ma liczne zagłębienia - bruzdy (sulci cerebri) i znajduje się między nimi zwoje (żyri cerebri). Kora mózgowa pokrywa całą powierzchnię zwojów i bruzd (stąd jej inna nazwa to paliusz - płaszcz), czasami wnikając głęboko w substancję mózgu.
Nasilenie i umiejscowienie bruzd i zwojów półkul mózgowych jest w pewnym stopniu zmienne, jednak główne powstają w procesie ontogenezy i są stałe, charakterystyczne dla każdego prawidłowo rozwiniętego mózgu.
14.2. GŁÓWNE GROOCHY I UCHWYTY PÓŁKUR MÓZGU
Górna boczna (wypukła) powierzchnia półkul (rys. 14.1a). Największy i najgłębszy boczny bruzda (bruzdy boczne),lub sylvian bruzda, - oddziela przednią i przednią część płata ciemieniowego od płata skroniowego znajdującego się poniżej. Płat czołowy i ciemieniowy są oddzielone centralny lub Roland, bruzda(bruzdy centralne), która przecina górną krawędź półkuli i biegnie w dół i do przodu wzdłuż jej wypukłej powierzchni, nieco krótszej od bocznego rowka. Płat ciemieniowy jest oddzielony od płata potylicznego znajdującego się za nim przez ciemieniowo-potyliczne i poprzeczne rowki potyliczne przechodzące wzdłuż przyśrodkowej powierzchni półkuli.
W płacie czołowym przed centralnym zakrętem i równolegle do niego znajduje się przedcentralny (zakręt precentralis), lub przedni centralny, zakręt, który jest ograniczony do przodu przez bruzdę przedcentralną (bruzda precentralis). Górne i dolne rowki czołowe odchodzą do przodu od bruzdy przedśrodkowej, dzieląc wypukłą powierzchnię przednich odcinków płata czołowego na trzy zakręty czołowe - górny, środkowy i dolny (gyri frontales superior, media i inferior).
Przednia część wypukłej powierzchni płata ciemieniowego znajduje się za bruzdą środkową postcentralną (zakręt postcentralis), lub tylny centralny, zakręt. Za nim graniczy z bruzdą postcentralną, od której rozciąga się bruzda śródciemieniowa. (bruzdy intraparietalis), oddzielanie górnych i dolnych płatków ciemieniowych (lobuli parietales superior i inferior). Z kolei w dolnym płatku ciemieniowym wyróżnia się zakręt nadbrzeżny (zakręt supramarginalis), otaczające tylną część bocznego (Sylvian) rowka i zakrętu kątowego (girus angularis), graniczący z tyłu górnego zakrętu skroniowego.
Na wypukłej powierzchni płata potylicznego mózgu bruzdy są płytkie i mogą się znacznie różnić, w wyniku czego zmienny jest również charakter zwojów znajdujących się między nimi.
Wypukła powierzchnia płata skroniowego jest podzielona przez górne i dolne bruzdy skroniowe, które są prawie równoległe do bruzdy bocznej (Sylvian), dzieląc wypukłą powierzchnię płata skroniowego na górny, środkowy i dolny zakręt skroniowy (gyri temporales superior, media et inferior). Górny zakręt skroniowy tworzy dolną wargę bruzdy bocznej (Sylvian). Na jego powierzchni skierowanej
po stronie bruzdy bocznej znajduje się kilka małych bruzd poprzecznych, podkreślających na niej mały zakręt poprzeczny (zakręt Geschla), co widać tylko po rozłożeniu brzegów bruzdy bocznej.
Przednia część bocznego (sylwiańskiego) rowka to zagłębienie z szerokim dnem, tworzące tzw wyspa (wyspa) lub płat wyspowy (lubus insularis). Górna krawędź bruzdy bocznej pokrywającej tę wyspę nazywa się opona (wieczko).
Wewnętrzna (środkowa) powierzchnia półkuli (Rys. 14.1b). Środkowa część wewnętrznej powierzchni półkuli jest ściśle powiązana ze strukturami międzymózgowia, od którego jest odgraniczona przez te związane z dużym mózgiem sklepienie (skrzydło) oraz Ciało modzelowate (Ciało modzelowate). Ten ostatni jest ograniczony z zewnątrz bruzdą ciała modzelowatego (sulcus corporis callosi), zaczynając od przodu - dziób (rostrum) i kończący się na pogrubionym tylnym końcu (splenium). Tutaj bruzda ciała modzelowatego przechodzi w głęboką bruzdę hipokampa (sulcus hippocampi), która wnika głęboko w substancję półkuli, wciskając ją do jamy dolnego rogu komory bocznej, w wyniku czego - powstaje róg amonowy.
Nieco odchodząc od bruzdy ciała modzelowatego i bruzdy hipokampa, znajduje się ciało modzelowate, bruzda podciemieniowa i nosowa, które są kontynuacją siebie. Te rowki wyznaczają od zewnątrz łukowatą część przyśrodkowej powierzchni półkuli mózgowej, znaną jako płat limbiczny(lobus limbicus). W płacie limbicznym występują dwa zwoje. Górna część płata limbicznego to górna część limbiczna (górna brzeżna) lub obręcz, zakręt (girus cinguli), dolną część tworzy dolny zakręt limbiczny lub zakręt konika morskiego (girus hipokamp), lub zakrętu przyhipokampowego (girus parahypocampalis), przed którym znajduje się haczyk (uncus).
Wokół płata limbicznego mózgu znajdują się formacje wewnętrznej powierzchni płatów czołowych, ciemieniowych, potylicznych i skroniowych. Większość wewnętrznej powierzchni płata czołowego zajmuje przyśrodkowa strona górnego zakrętu czołowego. Na granicy płatów czołowych i ciemieniowych półkuli mózgowej znajduje się płacik przyśrodkowy (lobulis paracentralis), co jest jakby kontynuacją przednich i tylnych zakrętów centralnych na przyśrodkowej powierzchni półkuli. Na granicy płatów ciemieniowych i potylicznych wyraźnie widoczna jest bruzda ciemieniowo-potyliczna. (bruzdy parietooccipitalis). Od dołu odchodzi z powrotem bruzda ostroga (bruzda calcarinus). Między tymi głębokimi bruzdami znajduje się trójkątny zakręt, zwany klinem. (klin). Przed klinem znajduje się czworokątny zakręt, związany z płatem ciemieniowym mózgu, przedklinkiem.
Dolna powierzchnia półkuli (Rys. 14.1c). Dolna powierzchnia półkuli mózgowej składa się z formacji płatów czołowych, skroniowych i potylicznych. Część płata czołowego przylegająca do linii środkowej to zakręt bezpośredni (girus rectus). Na zewnątrz jest ograniczony rowkiem węchowym (bruzdy węchowe), do których przylegają od dołu formacje analizatora węchowego: opuszka węchowa i droga węchowa. Bocznie od niego, aż do bocznego (Sylvian) rowka, który rozciąga się do dolnej powierzchni płata czołowego, znajdują się małe zakręty oczodołu (zakręt oczodołowy). Boczne odcinki dolnej powierzchni półkuli za bruzdą boczną zajmuje dolny zakręt skroniowy. Środkowy do niej jest boczny zakręt skroniowo-potyliczny. (gyrus occipitotemporalis lateralis), lub wrzecionowaty rowek. Zanim-
jego wewnętrzne części graniczą z zakrętem hipokampa, a tylne - z językiem (zakręt językowy) lub środkowy zakręt skroniowo-potyliczny (gyrus occipitotemporalis medialis). Ten ostatni, tylnym końcem, przylega do rowka ostrogi. Przednie odcinki zakrętów wrzecionowatych i językowych należą do płata skroniowego, a tylne do płata potylicznego mózgu.
14.3. BIAŁA MATERIA WIELKICH PÓŁKUR
Istota biała półkul mózgowych składa się z włókien nerwowych, głównie mieliny, które tworzą szlaki zapewniające połączenia między neuronami kory a skupiskami neuronów tworzących wzgórze, węzły podkorowe i jądra. Główna część istoty białej półkul mózgowych znajduje się w jej głębi środek półowalny, czyli promienna korona (korona promienista), składający się głównie z aferentnych i eferentnych występ drogi łączące korę mózgową z węzłami podkorowymi, jądrami i substancją siateczkową międzymózgowia i pnia mózgu, z odcinkami rdzenia kręgowego. Są one szczególnie zwarte pomiędzy wzgórzem a węzłami podkorowymi, gdzie tworzą wewnętrzną torebkę opisaną w rozdziale 3.
Nazywa się włókna nerwowe łączące części kory jednej półkuli asocjacyjny. Im krótsze włókna i połączenia, które tworzą, tym bardziej są powierzchowne; dłuższe połączenia asocjacyjne, położone głębiej, łączą stosunkowo odległe części kory mózgowej (ryc. 14.2 i 14.3).
Nazywa się włókna łączące półkule mózgowe i dlatego mają wspólną orientację poprzeczną komisaryczny, lub spanie. Włókna spoidłowe łączą identyczne części półkul mózgowych, stwarzając możliwość łączenia ich funkcji. Tworzą się trzy kolce duży mózg: najbardziej masywny z nich - Ciało modzelowate (Ciało modzelowate), dodatkowo składają się włókna spoidłowe spoidło przednie, znajduje się pod dziobem ciała modzelowatego (mównica corporis collosum) i łączącą zarówno regiony węchowe, jak i komisariat skarbca (commissura fornicis), lub spoidło hipokampa utworzone z włókien łączących struktury rogów amonu obu półkul.
W przedniej części ciała modzelowatego znajdują się włókna łączące płaty czołowe, następnie włókna łączące płaty ciemieniowe i skroniowe, tylna część ciała modzelowatego łączy płaty potyliczne mózgu. Spoidło przednie i spoidło sklepienia łączy głównie sekcje kory starożytnej i starej obu półkul, ponadto spoidło przednie zapewnia połączenie między ich środkowym i dolnym zakrętem skroniowym.
14.4. System węchowy
W procesie filogenezy rozwój dużego mózgu wiąże się z tworzeniem układu węchowego, którego funkcje przyczyniają się do zachowania żywotności zwierząt i mają niemałe znaczenie dla życia ludzkiego.
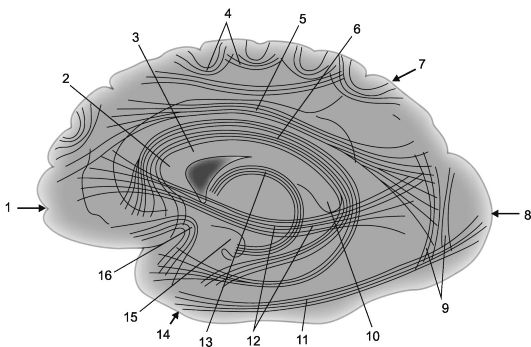
Ryż. 14.2.Asocjacyjne połączenia korowo-korowe w półkulach mózgowych [według V.P. Worobiow].
1 - płat czołowy; 2 - kolano ciała modzelowatego; 3 - ciało modzelowate; 4 - łukowate włókna; 5 - górna belka podłużna; 6 - zakręt obręczy; 7 - płat ciemieniowy, 8 - płat potyliczny; 9 - pionowe wiązki Wernickego; 10 - wałek ciała modzelowatego;
11 - dolna belka podłużna; 12 - wiązka podprzyczynowa (wiązka czołowo-potyliczna dolna); 13 - sklepienie; 14 - płat skroniowy; 15 - hak zakrętu hipokampu; 16 - wiązki haków (fasciculus uncinatus).
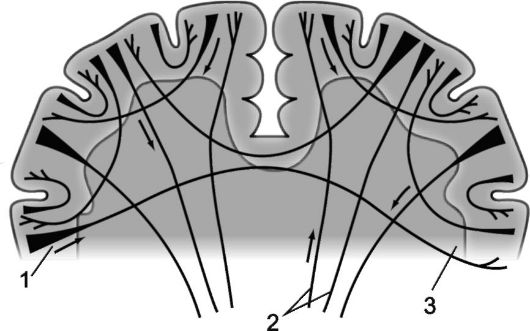
Ryż. 14.3.Mieloarchitektonika półkul mózgowych.
1 - włókna projekcyjne; 2 - włókna spoidłowe; 3 - włókna asocjacyjne.
14.4.1. Struktura układu węchowego
Ciała pierwszych neuronów układu węchowego znajdują się w błonie śluzowej głównie nos górna część przegrody nosowej i górnego kanału nosowego. Komórki węchowe są dwubiegunowe. Ich dendryty wychodzą na powierzchnię błony śluzowej i kończą się tutaj specyficznymi receptorami oraz aksony są zgrupowane w tzw filamenty węchowe (filiolfactorii), których liczba po każdej stronie wynosi około dwudziestu. Taki wiązka włókien węchowych i tworzy nerw czaszkowy I lub węchowy(rys. 14.4). Te wątki przejść do przedniego (węchowego, węchowego) dołu czaszki przez kość sitową i kończy się na komórki znajdujące się tutaj opuszki węchowe. Opuszki węchowe i proksymalne drogi węchowe są w rzeczywistości konsekwencją wypukłości materii dużego mózgu powstałej w procesie ontogenezy i reprezentują struktury z nią związane.
Opuszki węchowe zawierają komórki będące ciałami drugich neuronów. szlak węchowy, którego aksony tworzą się drogi węchowe (tracti olfactorii), znajduje się pod rowkami węchowymi, bocznie do bezpośrednich zwojów znajdujących się na podstawowej powierzchni płatów czołowych. Drogi węchowe są skierowane do tyłu do podkorowych ośrodków węchowych. Zbliżając się do przedniej płyty perforowanej, włókna dróg węchowych dzielą się na wiązki przyśrodkowe i boczne, tworząc z każdej strony trójkąt węchowy. Później te włókna są odpowiednie do ciał trzecich neuronów analizatora węchowego, znajdujących się

Ryż. 14.4.Analizator węchowy.
1 - komórki węchowe; 2 - nici węchowe (w sumie tworzą nerwy węchowe); 3 - opuszki węchowe; 4 - drogi węchowe; 5 - trójkąty węchowe; 6 - zakręt przyhipokampowy; 7 - strefa projekcyjna analizatora węchowego (schemat uproszczony).
w obszarze okrężnym i podmeluszkowym, w jądrach przeźroczystej przegrody, położonych przed spoidłem przednim. Spoidło przednie łączy oba regiony węchowe, a także zapewnia ich połączenie z układem limbicznym mózgu. Część aksonów trzeciego neuronu analizatora węchowego, przechodząca przez przednią spoidło mózgu, krzyżuje się.
Aksony trzeciego neuronu analizator węchowy, zlokalizowany w podkorowych ośrodkach węchowych, zmierzać w kierunku filogenetycznie stara skorupa przyśrodkowo-podstawna powierzchnia płata skroniowego (do zakrętu gruszkowatego i przyhipokampowego oraz do haczyka), gdzie znajduje się projekcyjna strefa węchowa, lub korowy koniec analizatora węchowego (pole 28, według Brodmana).
Układ węchowy jest więc jedynym układem czuciowym, w którym określone impulsy omijają wzgórze w drodze od receptorów do kory. Jednakże, Układ węchowy ma szczególnie wyraźne powiązania ze strukturami limbicznymi mózgu, a otrzymywane przez niego informacje mają istotny wpływ na stan sfery emocjonalnej i funkcje autonomicznego układu nerwowego. Zapachy mogą być przyjemne i nieprzyjemne, wpływają na apetyt, nastrój, mogą powodować różne reakcje wegetatywne, w szczególności nudności, wymioty.
14.4.2. Badanie zmysłu węchu i znaczenia jego zaburzeń w diagnostyce miejscowej
Podczas badania stanu węchu należy dowiedzieć się, czy pacjent pachnie, czy te odczucia są takie same po obu stronach, czy pacjent rozróżnia charakter odczuwanych zapachów, czy ma halucynacje węchowe - napadowe odczucia węchowe, które są nieobecne w środowisku.
Do badania węchu stosuje się substancje zapachowe, których zapach nie jest ostry (ostre zapachy mogą powodować podrażnienie receptorów nerwu trójdzielnego zlokalizowanych w błonie śluzowej nosa) i są znane pacjentowi (w przeciwnym razie trudno jest rozpoznać perwersja zapachu). Zmysł węchu jest sprawdzany z każdej strony osobno, drugie nozdrze musi być zamknięte. Możesz użyć specjalnie przygotowanych zestawów słabych roztworów substancji zapachowych (mięta, smoła, kamfora itp.), W praktyce można również zastosować środki improwizowane (chleb żytni, mydło, banan itp.).
Zmniejszony węch - hiposmia, brak zapachu - anosmia, podwyższony węch - hiperosmia, wypaczenie zapachów dysosmia, uczucie węchu przy braku bodźca - parosmia, subiektywne odczucie nieprzyjemnego zapachu, który faktycznie istnieje i jest spowodowany patologią organiczną w nosogardzieli - kakosmija, zapachy, które tak naprawdę nie istnieją, które pacjent odczuwa napadowo – halucynacje węchowe – są częściej aurą węchową padaczki skroniowej, co może być spowodowane rózne powody, w szczególności guz płata skroniowego.
Hiposmia lub anosmia po obu stronach jest zwykle wynikiem uszkodzenia błony śluzowej nosa w wyniku ostrego kataru, grypy, alergicznego nieżytu nosa, zaniku błony śluzowej
nosa z powodu przewlekłego nieżytu nosa i długotrwałego stosowania kropli do nosa zwężających naczynia. Przewlekły nieżyt nosa z zanikiem błony śluzowej nosa (zanikowy nieżyt nosa), choroba Sjögrena skazuje człowieka na uporczywą anosmię. Obustronna hiposmia może być spowodowana niedoczynnością tarczycy, cukrzycą, hipogonadyzmem, niewydolnością nerek, długotrwałym kontaktem z metalami ciężkimi, formaldehydem itp.
Jednakże, jednostronna hiposmia lub anosmia jest często wynikiem guza wewnątrzczaszkowego, częściej oponiaka przedniego dołu czaszki (węchowego), co stanowi do 10% oponiaków śródczaszkowych, a także niektórych guzów glejowych płata czołowego. Zaburzenia węchowe powstają w wyniku ucisku drogi węchowej po stronie ogniska patologicznego i przez pewien czas mogą być jedynym ogniskowym objawem choroby. Nowotwory można wizualizować za pomocą skanowania CT lub MRI. Wraz ze wzrostem oponiaka dołu węchowego z reguły rozwijają się zaburzenia psychiczne charakterystyczne dla zespołu czołowego (patrz rozdział 15).
Jednostronne uszkodzenie części analizatora węchowego znajdujących się nad jego ośrodkami podkorowymi, z powodu niepełnego odkupienia dróg na poziomie przedniej spoidła mózgowego, zwykle nie prowadzi do znacznego zmniejszenia węchu. Podrażnienie przez patologiczny proces kory części przyśrodkowo-podstawnych płata skroniowego, głównie zakrętu hipokampa i jego haczyka, może powodować napadowe wystąpienie halucynacje węchowe. Pacjent nagle zaczyna pachnieć bez powodu, często o nieprzyjemnym charakterze (zapach spalonych, zgniłych, zgniłych, spalonych itp.). Halucynacje węchowe w obecności ogniska epileptogennego w okolicy przyśrodkowej płata skroniowego mózgu może być przejawem aury napadu padaczkowego. Klęska części proksymalnej, w szczególności korowego końca analizatora węchowego, może powodować umiarkowaną obustronną (bardziej po przeciwnej stronie) hiposmię i upośledzoną zdolność do identyfikacji i różnicowania zapachów (agnozja węchowa). Ostatnia postać zaburzenia węchowego, która objawia się w starszym wieku, najprawdopodobniej wiąże się z naruszeniem funkcji kory w wyniku procesów zanikowych w jej projekcyjnej strefie węchowej.
14.5. KOMPLEKS LIMBIC-SIATKÓWKI
W 1878 r. P. Broca(Broca P., 1824-1880) pod nazwą „duży płat brzeżny lub limbiczny” (od łac. limbus - edge) łączył hipokamp i zakręt obręczy, połączone za pomocą przesmyku zakrętu obręczy, znajdującego się nad grzbietem ciała modzelowatego.
W 1937 D. Papets(Papez J.), na podstawie danych eksperymentalnych, wysunęli uzasadniony sprzeciw wobec dotychczasowej koncepcji zaangażowania struktur przypodstawnych półkul mózgowych głównie w dostarczanie węchu. On zasugerowali, że główna część przyśrodkowej części półkuli mózgowej, zwanej wówczas mózgiem węchowym (rhinencephalon), do którego należy płat limbiczny, stanowi morfologiczną podstawę nerwowego mechanizmu zachowań afektywnych i połączyła je pod nazwą„emocjonalny krąg” który obejmował podwzgórze,
przednie jądra wzgórza, zakręt obręczy, hipokamp i ich połączenia. Od tego czasu struktury te są również określane przez fizjologów jako wokół Papetza.
pojęcie "mózg trzewny" zasugerował P.D. McLean (1949), oznaczający w ten sposób złożony związek anatomiczno-fizjologiczny, który od 1952 roku nazywa się „układ limbiczny”. Później okazało się, że układ limbiczny bierze udział w pełnieniu różnych funkcji, a obecnie większość z nich, w tym zakręt obręczy i hipokamp (parahipokamp) jest zwykle łączony w obszar limbiczny, który ma liczne powiązania ze strukturami układu limbicznego. formacja siatkowata, nadrabiająca to kompleks limbiczno-siatkowy szerokie koło procesy fizjologiczne i psychologiczne.
Obecnie do płat limbiczny zwyczajowo przypisuje się elementy starej kory (archiocortex), obejmujące zakręt zębaty i zakręt hipokampa; starożytna kora (paleocortex) przedniego hipokampa; jak również środkowa lub pośrednia kora (mezokorteks) zakrętu obręczy. Termin „układ limbiczny” obejmuje elementy płata limbicznego i struktur pokrewnych - śródwęchowego (zajmującego większość zakrętu przyhipokampowego) i przegrody, a także kompleksu ciała migdałowatego i ciała wyrostka sutkowatego (Duus P., 1995).
Ciało wyrostka sutkowatego łączy struktury tego układu ze śródmózgowiem i formacją siatkowatą. Impulsy pochodzące z układu limbicznego mogą być przekazywane przez przednie jądro wzgórza do zakrętu obręczy i do kory nowej drogami utworzonymi przez włókna asocjacyjne. Impulsy pochodzące z podwzgórza mogą dotrzeć do kory oczodołowo-czołowej i przyśrodkowego jądra grzbietowego wzgórza.
Liczne bezpośrednie i informacja zwrotna zapewniają wzajemne połączenie i współzależność struktur limbicznych i wielu formacji międzymózgowia i ustnych części pnia mózgu (niespecyficzne jądra wzgórza, podwzgórza, skorupy, wędzidełka, tworzenie siateczkowate pnia mózgu), a także z jądrami podkorowymi (blade, skorupa, jądra ogoniastego) oraz z nową korą mózgową półkul mózgowych, głównie z korą płatów skroniowych i czołowych.
Pomimo różnic filogenetycznych, morfologicznych i cytoarchitektonicznych wiele z wymienionych struktur (obszar limbiczny, struktury centralne i przyśrodkowe wzgórza, podwzgórze, tworzenie siateczkowate pnia mózgu) zalicza się zazwyczaj do tzw. kompleks limbiczno-siatkowy, która działa jako strefa integracji wielu funkcji, zapewniając organizację polimodalnych, holistycznych reakcji organizmu na różne wpływy, co jest szczególnie widoczne w sytuacjach stresowych.
Struktury kompleksu limbiczno-siatkowego mają duża liczba wejścia i wyjścia, przez które przechodzą błędne kręgi licznych połączeń aferentnych i eferentnych, zapewniających połączone funkcjonowanie formacji wchodzących w skład tego kompleksu i ich interakcja ze wszystkimi częściami mózgu, w tym z korą mózgową.
W strukturach kompleksu limbiczno-siatkowego dochodzi do zbieżności wrażliwych impulsów, które występują w intero- i zewnętrzoreceptorach, w tym w polach receptorowych narządów zmysłów. Na tej podstawie w kompleksie limbiczno-siatkowym pierwotna synteza informacji o stanie wewnętrznego środowiska organizmu, a także o czynnikach wpływających na organizm otoczenie zewnętrzne, kształtują się potrzeby elementarne, motywacje biologiczne i towarzyszące im emocje.
Kompleks limbiczno-siatkowy określa stan sfery emocjonalnej, uczestniczy w regulacji relacji wegetatywno-trzewnych mających na celu utrzymanie względnej stałości środowiska wewnętrznego (homeostazy), a także dostaw energii i korelacji czynności ruchowych. Poziom świadomości, możliwość zautomatyzowanych ruchów, aktywność funkcji motorycznych i umysłowych, mowa, uwaga, zdolność orientacji, pamięć, zmiana czuwania i snu zależą od jej stanu.
Uszkodzeniu struktur kompleksu limbiczno-siatkowego mogą towarzyszyć różnorodne objawy kliniczne: wyraźne zmiany w sferze emocjonalnej o charakterze trwałym i napadowym, anoreksja lub bulimia, zaburzenia seksualne, zaburzenia pamięci, w szczególności objawy zespołu Korsakoffa, w którym pacjent traci zdolność zapamiętywania bieżących zdarzeń (zachowuje bieżące zdarzenia w pamięci nie dłużej niż 2 minuty), zaburzenia autonomiczno-endokrynologiczne, zaburzenia snu, zaburzenia psychosensoryczne w postaci złudzeń i halucynacji, zmiany świadomości, kliniczne objawy mutyzmu akinetycznego, napady padaczkowe.
Do tej pory przeprowadzono wiele badań dotyczących badania morfologii, relacji anatomicznych, funkcji regionu limbicznego i innych struktur wchodzących w skład kompleksu limbiczno-siatkowego, jednakże fizjologii i cech obrazu klinicznego jego zmiany dziś nadal w dużej mierze wymagają wyjaśnienia. Większość informacji o jego funkcji, zwłaszcza funkcje obszaru przyhipokampowego, uzyskane w doświadczeniach na zwierzętach metody podrażnienia, ekstyrpacji lub stereotaksji. Uzyskane w ten sposób wyniki wymagają ostrożności przy ekstrapolacji na ludzi. Specjalne znaczenie prowadzić obserwacje kliniczne pacjentów z uszkodzeniami obszarów przyśrodkowej półkuli mózgowej.
W latach 50. i 60. XX wieku. w trakcie rozwoju psychochirurgii pojawiły się doniesienia o leczeniu pacjentów z nieuleczalną zaburzenia psychiczne i przewlekłego zespołu bólowego poprzez obustronną cingulotomię (rozwarstwienie zakrętu obręczy), przy czym zwykle stwierdzano regresję lęku, stany obsesyjne, pobudzenie psychoruchowe, zespoły bólowe, co uznano za dowód udziału zakrętu obręczy w tworzeniu emocji i ból. Jednocześnie biningulotomia prowadziła do głębokich zaburzeń osobowości, dezorientacji, zmniejszenia krytyczności stanu i euforii.
W monografii N.N. Bragina (1974). Autor dochodzi do wniosku, że zespół skroniowo-podstawny obejmuje zaburzenia trzewno-wegetatywne, ruchowe i psychiczne, zwykle objawiające się kompleksowo. Cała różnorodność objawów klinicznych N.N. Bragin sprowadza się do dwóch głównych wieloczynnikowych wariantów patologii z przewagą zjawisk „podrażniających” i „hamujących”.
Pierwsza z nich to zaburzenia emocjonalne z towarzyszącym niepokojem ruchowym (wzrost pobudliwości, gadatliwości, nerwowość, uczucie wewnętrznego niepokoju), napady lęku, udręka życiowa, różne zaburzenia trzewno-wegetatywne (zmiany tętna, oddychania, zaburzenia żołądkowo-jelitowe, gorączka, wzmożona potliwość itp.). U tych pacjentów, na tle ciągłego niepokoju ruchowego, często dochodziło do ataków pobudzenia motorycznego.
Nija. EEG tej grupy pacjentów charakteryzowało się łagodnymi zmianami mózgowymi w kierunku integracji (przyspieszony i zaostrzony rytm alfa, rozproszone oscylacje beta). Powtarzane bodźce doprowadzające wywoływały wyraźne odpowiedzi EEG, które w przeciwieństwie do normalnych nie zanikały w miarę powtarzania bodźców.
Druga („hamująca”) odmiana zespołu przyśrodkowego charakteryzuje się zaburzeniami emocjonalnymi w postaci depresji z upośledzeniem ruchowym (obniżony nastrój, zubożenie i spowolnienie tempa procesów psychicznych, zmiany zdolności motorycznych, przypominające zespół akinetyczno-sztywny Mniej charakterystyczne są napady trzewno-wegetatywne odnotowane w pierwszej grupie.W EEG pacjentów z tej grupy charakteryzowały się zmiany mózgowe, objawiające się przewagą wolnych form aktywności (nieregularny, powolny rytm alfa, grupy oscylacji theta, rozlane fale delta ). Uwagę przyciągnął ostry spadek reaktywności EEG.
Pomiędzy tymi dwoma skrajnymi wariantami występowały również pośrednie z przejściowymi i mieszanymi kombinacjami poszczególnych objawów. Niektóre z nich charakteryzują się więc stosunkowo słabymi objawami pobudzenia depresyjnego z wzmożoną aktywnością ruchową i zmęczeniem, z przewagą wrażeń seneopatycznych, podejrzliwości, która u części pacjentów dochodzi do stanów paranoidalnych oraz majaczenia hipochondrycznego. Drugą grupę pośrednią wyróżniało ekstremalne nasilenie objawów depresyjnych na tle sztywności pacjenta.
Dane te pozwalają nam mówić o podwójnym (aktywującym i hamującym) wpływie hipokampu i innych struktur obszaru limbicznego na reakcje behawioralne, emocje, stan psychiczny i aktywność bioelektryczną kory. Obecnie złożone zespoły kliniczne tego typu nie powinny być uważane za ognisko pierwotne. Raczej należy je rozpatrywać w świetle pomysłów dotyczących system wielopoziomowy organizacja aktywności mózgu.
S.B. Buklina (1997) przytoczył dane z badania 41 pacjentów z malformacjami tętniczo-żylnymi w okolicy zakrętu obręczy. Przed operacją 38 pacjentów miało zaburzenia pamięci na czele, a 5 z nich miało objawy zespołu Korsakoffa, u 3 pacjentów zespół Korsakoffa pojawił się po zabiegu, natomiast nasilenie nasilenia wad pamięci korelowało ze stopniem zniszczenia zakrętu obręczy sam, a także z udziałem w procesie patologicznym sąsiednich struktur ciała modzelowatego, podczas gdy zespół amnezyjny nie zależał od strony lokalizacji wady i jej lokalizacji na długości zakrętu obręczy.
Głównymi cechami rozpoznanych zespołów amnestycznych były zaburzenia odtwarzania bodźców słuchowo-mowy, naruszenia selektywności śladów w postaci wtrąceń i zanieczyszczeń oraz niemożność zachowania znaczenia w przekazywaniu opowieści. U większości pacjentów krytyczność oceny ich stanu była zmniejszona. Autor zwrócił uwagę na podobieństwo tych zaburzeń do ubytków amnestycznych u pacjentów ze zmianami czołowymi, co można tłumaczyć obecnością połączeń między zakrętem obręczy a płatem czołowym.
Więcej rozległe procesy patologiczne w obszarze limbicznym powodują wyraźne zaburzenia funkcji wegetatywno-trzewnych.
Ciało modzelowate(Ciało modzelowate)- największa spoidła między półkulami mózgowymi. Jego przednie części, w szczególności kolano ciała modzelowatego
ciało (rodzaj corporis callosi), połączyć płaty czołowe, sekcje środkowe - pień ciała modzelowatego (truncus corporis callosi)- zapewniają komunikację między odcinkami skroniowymi i ciemieniowymi półkul, odcinkami tylnymi, w szczególności grzbietem ciała modzelowatego (splenium corporis callosi), połączyć płaty potyliczne.
Uszkodzeniom ciała modzelowatego zwykle towarzyszą zaburzenia stanu psychicznego pacjenta. Zniszczenie jego przedniej części prowadzi do rozwoju „czołowej psychiki” (spontaniczność, naruszenie planu działania, zachowanie, krytyka, charakterystyczne dla czołowy zespół zrogowaciały - akinezja, amimia, aspontaniczność, astazja-abazja, apraksja, chwytanie odruchów, demencja). Odłączenie połączeń między płatami ciemieniowymi prowadzi do perwersji zrozumienie "plany ciała" oraz pojawienie się apraksji głównie w lewej ręce. Dysocjacja płatów skroniowych może się zamanifestować naruszenie percepcji środowiska zewnętrznego, utrata prawidłowej orientacji w nim (zaburzenia amnestyczne, konfabulacje, syndrom tego, co już widziane itp.). Ogniska patologiczne w tylnych częściach ciała modzelowatego często charakteryzują się oznakami agnozji wzrokowej.
14.6. ARCHITEKTONIKA KORY MÓZGU
Struktura kory mózgowej jest niejednorodna. Mniej złożona w budowie, wcześnie powstająca w procesie filogenezy starożytna kora (archiocortex) i stara kora (paleokorteks), związane z przeważnie do płata limbicznego mózg. Większa część kory mózgowej (95,6%), ze względu na jej późniejsze ukształtowanie filogenetyczne, nosi nazwę nowa kora (neocortex) i ma znacznie bardziej złożoną strukturę wielowarstwową, ale też niejednorodną w różnych jej strefach.
W związku z faktem, że architektura kory jest w pewnym związku z jej funkcją, wiele uwagi poświęcono jej badaniu. Jednym z twórców doktryny cytoarchitektoniki kory mózgowej był V.A. Betz (1834-1894), który po raz pierwszy w 1874 r. opisał duże komórki piramidalne kory ruchowej (komórki Betza) i określił zasady podziału kory mózgowej na główne obszary. W przyszłości wielki wkład w rozwój teorii budowy kory wniosło wielu badaczy - A. Campbell (A. Cambell), E. Smith (E. Smith), K. Brodmann (K. Brodmann) ), Oscar Vogt i Cecilia Vogt (O. Vogt, S. Vogt). Ogromne zasługi w badaniu architektury kory mózgowej mają pracownicy Instytutu Mózgu Akademii Nauk Medycznych (S.A. Sarkisov, N.I. Filimonov, E.P. Kononova itp.).
Główny typ struktury nowej skorupy (Rys. 14.5), z którym porównywane są wszystkie jego sekcje - kora składająca się z 6 warstw (kora homotypowa, według Brodmana).
Warstwa I - molekularna lub strefowa, najbardziej powierzchowna, uboga w komórki, jej włókna mają kierunek, głównie równoległy do powierzchni kory.
II warstwa - zewnętrzna ziarnista. Składa się z dużej liczby gęsto ułożonych małych ziarnistych komórek nerwowych.
III warstwa - piramidy małe i średnie, najszersze. Składa się z komórek piramidalnych, których rozmiary nie są takie same, co pozwala podzielić tę warstwę na podwarstwy w większości pól korowych.
Warstwa IV - ziarnista wewnętrzna. Składa się z gęsto ułożonych drobnych komórek-ziaren o okrągłym i kanciastym kształcie. Ta warstwa jest najbardziej zmienna
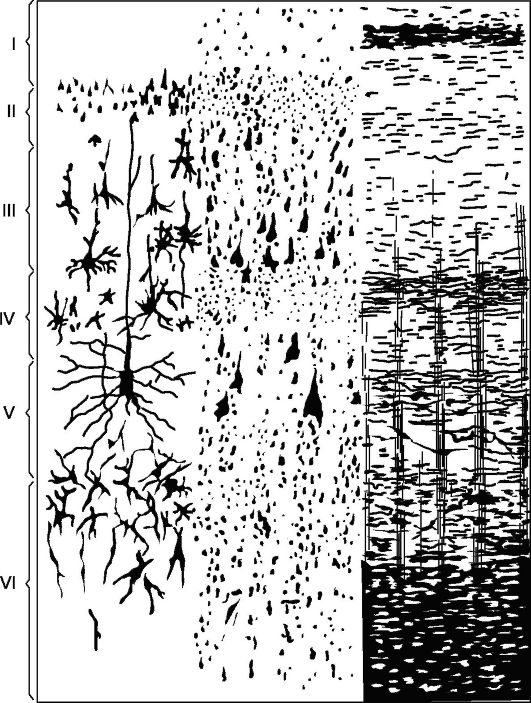
Ryż. 14.5.Cytoarchitektonika i mieloarchitektonika strefy ruchowej kory mózgowej.
Po lewej: I - warstwa molekularna; II - zewnętrzna warstwa ziarnista; III - warstwa małych i średnich piramid; IV - wewnętrzna warstwa ziarnista; V - warstwa dużych piramid; VI - warstwa komórek polimorficznych; po prawej - elementy mieloarchitektoniki.
na niektórych polach (na przykład pole 17) dzieli się na podwarstwy, w niektórych miejscach gwałtownie staje się cieńszy, a nawet całkowicie zanika.
Warstwa V - duże piramidy lub zwojowe. Zawiera duże komórki piramidalne. W niektórych obszarach mózgu warstwa jest podzielona na podwarstwy, w strefie motorycznej składa się z trzech podwarstw, z których środkowa zawiera gigantyczne komórki piramidalne Betza, osiągające średnicę 120 mikronów.
Warstwa VI - komórki polimorficzne lub wielopostaciowe. Składa się głównie z ogniw w kształcie trójkątnego wrzeciona.
Struktura kory mózgowej ma dużą liczbę zmienności ze względu na zmiany grubości poszczególnych warstw, przerzedzenie lub zanik lub,
przeciwnie, pogrubienie i podział na podwarstwy niektórych z nich (strefy heterotypowe według Brodmana).
Kora każdej półkuli mózgu jest podzielona na kilka regionów: potyliczny, górny i dolny ciemieniowy, postcentralny, centralny, przedcentralny, czołowy, skroniowy, limbiczny, wyspowy. Każda z nich zgodnie z charakterystyką podzielone na kilka pól, ponadto każde pole ma swoje własne konwencjonalne oznaczenie porządkowe (ryc. 14.6).
Badania nad architekturą kory mózgowej, wraz z badaniami fizjologicznymi, w tym elektrofizjologicznymi, badaniami i obserwacjami klinicznymi w znacznym stopniu przyczyniły się do rozwiązania problemu rozkładu funkcji w korze mózgowej.
14.7. POLA PROJEKCJI I STOWARZYSZENIA CORTUS
W procesie opracowywania doktryny o roli kory mózgowej i jej poszczególnych odcinków w pełnieniu określonych funkcji pojawiały się różne, niekiedy przeciwstawne punkty widzenia. Tak więc istniała opinia o ściśle lokalnej reprezentacji w korze mózgowej wszystkich ludzkich zdolności i funkcji, aż do najbardziej złożonych, umysłowych (lokalizacja, psychomorfologizm). Sprzeciwiła mu się inna opinia o absolutnej funkcjonalnej równoważności wszystkich części kory mózgowej (ekwipotencjalność).
Ważny wkład do teorii lokalizacji funkcji w korze mózgowej wniósł I.P. Pawłow (1848-1936). Wyróżnił strefy projekcyjne kory (korowe końce analizatorów). pewne rodzaje wrażliwość) i strefy asocjacyjne znajdujące się między nimi, badali procesy hamowania i wzbudzania w mózgu, ich wpływ na stan funkcjonalny kory mózgowej. Podział kory na strefy projekcyjne i asocjacyjne przyczynia się do zrozumienia organizacji pracy kory mózgowej i uzasadnia rozwiązywanie praktycznych problemów, w szczególności w diagnostyce miejscowej.
strefy projekcji zapewniają głównie proste specyficzne czynności fizjologiczne, przede wszystkim percepcję wrażeń o określonej modalności. Zbliżające się do nich ścieżki projekcyjne łączą te strefy z terytoriami receptorów na peryferiach, które pozostają z nimi w funkcjonalnej korespondencji. Przykładami projekcyjnych stref korowych są obszar tylnego centralnego zakrętu opisany już w poprzednich rozdziałach (strefa typy ogólne wrażliwość) lub obszar rowka ostrogi znajdującego się po przyśrodkowej stronie płata potylicznego (strefa wizualna projekcyjna).
Strefy stowarzyszeniowe kora nie ma bezpośrednich połączeń z obwodem. Znajdują się one pomiędzy strefami projekcyjnymi i posiadają liczne powiązania skojarzeniowe z tymi strefami projekcyjnymi oraz z innymi strefami skojarzeniowymi. Funkcją stref asocjacyjnych jest przeprowadzenie wyższej analizy i syntezy wielu elementarnych i bardziej złożonych składników. Tutaj w istocie istnieje zrozumienie informacji wchodzących do mózgu, powstawanie pomysłów i pojęć.
ŻOŁNIERZ AMERYKAŃSKI. Polyakov w 1969, na podstawie porównania architektury ludzkiej kory mózgowej i niektórych zwierząt, stwierdził, że asocjacyjne
Ryż. 14.6.Pola architektoniczne kory mózgowej [według Brodmana]. a - powierzchnia zewnętrzna; b - powierzchnia przyśrodkowa.
strefy w ludzkiej korze mózgowej wynoszą 50%, w korze małp wyższych (humanoidalnych) - 20%, u niższych małp liczba ta wynosi 10% (ryc. 14.7). Wśród obszarów asocjacyjnych kory ludzkiego mózgu, ten sam autor zasugerował izolację pola drugorzędne i trzeciorzędne. Drugorzędne pola asocjacyjne sąsiadują z polami rzutowymi. Dokonują analizy i syntezy elementarnych wrażeń, które wciąż zachowują określoną orientację.
Trzeciorzędne pola asocjacyjne leżą głównie pomiędzy drugorzędnymi i są nakładającymi się strefami sąsiednich terytoriów. Związane są one przede wszystkim z analityczną aktywnością kory, zapewniającą najwyższe funkcje umysłowe tkwiące w człowieku w jego najbardziej złożonych przejawach intelektualnych i mowy. Dojrzałość funkcjonalna trzeciego stopnia jako-
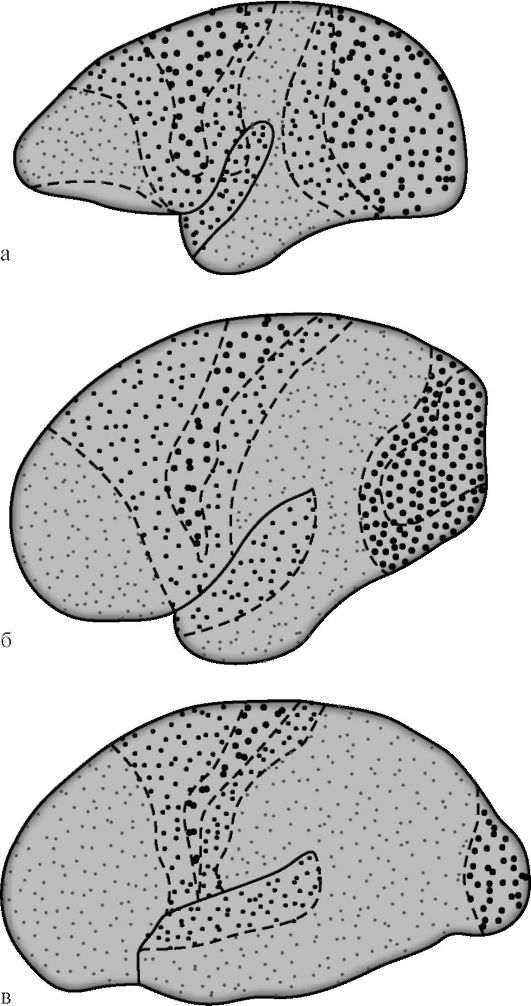
Ryż. 14.7. Zróżnicowanie projekcji i obszarów asocjacyjnych kory mózgowej podczas ewolucji naczelnych [według G.I. Poliakow]. a - mózg dolnej małpy; b - mózg wyższej małpy; c - ludzki mózg. Duże kropki oznaczają strefy projekcji, małe kropki - skojarzone. U niższych małp strefy asocjacyjne zajmują 10% powierzchni kory, u wyższych - 20%, u ludzi - 50%.
pola społeczne kory mózgowej występują najbardziej późno i tylko w sprzyjającym środowisku społecznym. W przeciwieństwie do innych pól korowych, trzeciorzędowe pola prawej i lewej półkuli charakteryzują się wyraźnym asymetria funkcjonalna.
14.8. MIEJSCOWE ROZPOZNANIE ZMIAN W KORCIE MÓZGU
14.8.1. Manifestacje uszkodzenia stref projekcyjnych kory mózgowej
W korze każdej półkuli mózgu, za zakrętem centralnym, znajduje się 6 stref projekcyjnych.
1. W przedniej części płata ciemieniowego, w rejonie tylnego centralnego zakrętu (pola cytoarchitektoniczne 1, 2, 3) usytuowany strefa projekcji ogólnych typów wrażliwości(rys. 14.4). Znajdujące się tutaj obszary kory otrzymują wrażliwe impulsy przychodzące wzdłuż ścieżek projekcyjnych o ogólnych typach wrażliwości z aparatu receptorowego przeciwnej połowy ciała. Im wyższy obszar tej strefy projekcyjnej kory, tym niżej położone części przeciwległej połowy ciała mają połączenia projekcyjne. Części ciała o rozległym odbiorze (język, powierzchnia dłoniowa dłoni) odpowiadają nieodpowiednio dużym obszarom obszaru stref projekcyjnych, podczas gdy pozostałe części ciała (kończyny bliższe, tułów) mają niewielką powierzchnię korową reprezentacja.
Podrażnienie przez patologiczny proces strefy korowej ogólnych typów wrażliwości prowadzi do ataku parestezji w częściach ciała odpowiadających podrażnionym obszarom kory mózgowej (wrażliwy napad Jacksona), który może przekształcić się w wtórnie uogólniony napad. Klęska końca korowego analizatora ogólnych typów wrażliwości może powodować rozwój hipopalgezji lub znieczulenia w odpowiedniej strefie przeciwnej połowy ciała, podczas gdy miejsce hipoestezji lub znieczulenia może mieć pionowe krążenie lub korzenie- typ segmentowy. W pierwszym przypadku zaburzenie wrażliwości objawia się po stronie przeciwnej do ogniska patologicznego w okolicy warg, kciuka lub w dystalnej części kończyny z okrężną obwódką, czasami jak skarpetka lub rękawiczka. W drugim przypadku strefa zaburzeń wrażliwości ma postać paska i znajduje się wzdłuż wewnętrznej lub zewnętrznej krawędzi ręki lub nogi; tłumaczy się to tym, że wewnętrzna strona kończyn jest prezentowana w przedniej, a zewnętrzna - w tylnych odcinkach strefy projekcji analizatora ogólnych typów wrażliwości.
2. Strefa projekcji wizualnej usytuowany w korze przyśrodkowej powierzchni płata potylicznego w okolicy bruzdy ostrogi (pole 17). Na tym polu następuje rozwarstwienie IV (wewnętrznej ziarnistej) warstwy kory z wiązką włókien mielinowych na dwie podwarstwy. Oddzielne sekcje pola 17 otrzymują impulsy z pewnych sekcji homonimicznych połówek siatkówek obu oczu; podczas gdy impulsy pochodzące z dolnych części homonimicznych połówek siatkówki docierają
kory dolnej wargi bruzdy ostrogi, a impulsy pochodzące z górnych części siatkówki kierowane są do kory jej górnej wargi.
Klęska patologicznego procesu strefy projekcji wzrokowej prowadzi do pojawienia się po przeciwnej stronie kwadrantu lub całkowitej homonimicznej hemianopii po stronie przeciwnej do ogniska patologicznego. Obustronne uszkodzenie pól korowych 17 lub prowadzących do nich projekcyjnych dróg wzrokowych może prowadzić do całkowitej ślepoty. Podrażnienie kory w strefie projekcji wzrokowej może powodować pojawienie się halucynacji wzrokowych w postaci fotopsji w odpowiednich częściach przeciwległych połówek pól widzenia.
3. Obszar projekcji słuchu usytuowany w korze zwojów Heschla na dolnej wardze bruzdy bocznej (Sylvian) (pola 41 i 42), które w rzeczywistości są częścią wyższego zakrętu skroniowego. Podrażnienie tej strefy kory może powodować występowanie halucynacji słuchowych (ataki odczuwania hałasu, dzwonienia, gwizdu, brzęczenia itp.). Zniszczenie strefy projekcji słuchowej z jednej strony może spowodować pewien ubytek słuchu w obu uszach, w większym stopniu w stosunku do ogniska patologicznego.
4 i 5. Strefy projekcji zapachowej i smakowej są na przyśrodkowej powierzchni sklepionego zakrętu (obszar limbiczny) mózgu. Pierwszy z nich znajduje się w zakręcie przyhipokampowym (pole 28). Strefa projekcyjna smaku jest zwykle zlokalizowana w korze okolicy wieczkowej (pole 43). Podrażnienie stref projekcji zapachu i smaku może powodować ich wypaczenie lub prowadzić do rozwoju odpowiednich halucynacji węchowych i smakowych. Jednostronna utrata funkcji stref projekcyjnych zapachu i smaku może spowodować nieznaczny spadek odpowiednio zapachu i smaku po obu stronach. Obustronne zniszczenie końców korowych tych samych analizatorów objawia się odpowiednio brakiem zapachu i smaku po obu stronach.
6. Strefa projekcji przedsionkowej. Jego lokalizacja nie jest określona. Jednocześnie wiadomo, że aparat przedsionkowy ma liczne połączenia anatomiczne i funkcjonalne. Możliwe, że lokalizacja reprezentacji układu przedsionkowego w korze nie została jeszcze wyjaśniona, ponieważ jest wieloogniskowa. N.S. Blagoveshchenskaya (1981) uważa, że w korze mózgowej strefy projekcji przedsionkowej są reprezentowane przez kilka anatomicznych i funkcjonalnych oddziałujących kompleksów, które znajdują się w polu 8, na styku płatów czołowych, skroniowych i ciemieniowych oraz w strefie zakrętów środkowych , natomiast zakłada się, że każdy z tych obszarów kory spełnia swoje własne funkcje. Pole 8 jest arbitralnym centrum spojrzenia, jego podrażnienie powoduje odwrócenie wzroku w kierunku przeciwnym do ogniska patologicznego, zmiany rytmu i charakteru oczopląsu doświadczalnego, szczególnie w krótkim czasie po napadzie padaczkowym. W korze płata skroniowego znajdują się struktury, których podrażnienie powoduje zawroty głowy, co objawia się w szczególności padaczką płata skroniowego; porażka obszarów reprezentacji struktur przedsionkowych w korze zakrętu centralnego wpływa na stan napięcia mięśni poprzecznie prążkowanych. Obserwacje kliniczne sugerują, że drogi przedsionkowo-jądrowo-korowe wykonują częściowe odkurzenie.
Należy podkreślić, że oznaki podrażnienia wymienionych stref projekcyjnych mogą być przejawem aury napadu padaczkowego o podobnym charakterze.
IP Pawłow uważał, że możliwe jest rozważenie kory przedśrodkowego zakrętu, która wpływa na funkcje motoryczne i napięcie mięśniowe głównie przeciwnej połowy ciała, z którą jest połączona głównie drogami korowo-jądrowymi i korowo-rdzeniowymi (piramidowymi), jako strefa projekcyjna tzw analizator silnika. Ta strefa zajmuje przede wszystkim pole 4, na które w odwróconej formie rzutowana jest przeciwległa połowa ciała. To pole zawiera większość gigantycznych komórek piramidalnych (komórek Betza), których aksony stanowią 2-2,5% wszystkich włókien ścieżki piramidalnej, a także średnie i małe komórki piramidalne, które wraz z aksonami tego samego Komórki znajdujące się w sąsiadującym z polem 4 bardziej rozległym polu 6, biorą udział w realizacji połączeń monosynaptycznych i polisynaptycznych korowo-mięśniowych. Połączenia monosynaptyczne zapewniają przede wszystkim szybkie i precyzyjne działania celowe, zależne od skurczów poszczególnych mięśni poprzecznie prążkowanych.
Uszkodzenie dolnych części strefy motorycznej zwykle prowadzi do rozwoju po przeciwnej stronie ramienno-twarzowy (ramiona twarzy) zespół lub zespół językowo-twarzowo-ramię, które często obserwuje się u pacjentów z zaburzeniami krążenia mózgowego w dorzeczu tętnicy środkowej mózgu, przy jednoczesnym niedowładzie mięśni twarzy, języka i ramienia, zwłaszcza barku w typie centralnym.
Podrażnienie kory strefy ruchowej (pola 4 i 6) prowadzi do pojawienia się drgawek w mięśniach rzutowanych na tę strefę lub grupy mięśniowe. Częściej są to drgawki miejscowe typu padaczki Jacksona, które mogą przekształcić się we wtórnie uogólniony napad padaczkowy.
14.8.2. Manifestacje uszkodzenia pól asocjacyjnych kory mózgowej
Pomiędzy strefami projekcyjnymi kory znajdują się pola asocjacyjne. Otrzymują impulsy głównie z komórek stref projekcyjnych kory. W polach asocjacyjnych odbywa się analiza i synteza informacji, które przeszły pierwotne przetwarzanie w polach projekcyjnych. Strefy asocjacyjne kory górnego płata ciemieniowego zapewniają syntezę elementarnych wrażeń, w związku z tym tak złożone typy wrażliwości, jak poczucie lokalizacji, poczucie wagi, poczucie dwuwymiarowo-przestrzenne, a także złożone powstają tu wrażenia kinestetyczne.
W rejonie bruzdy międzyciemieniowej znajduje się strefa asocjacyjna, która zapewnia syntezę wrażeń emanujących z części własnego ciała. Uszkodzenie tego regionu kory prowadzi do autopagnozja, tych. do nierozpoznawania lub ignorowania części własnego ciała lub do pseudomelia uczucie posiadania dodatkowej ręki lub nogi oraz anosognozja - brak świadomości wady fizycznej powstałej w związku z chorobą (np. paraliż lub niedowład kończyny). Zwykle wszystkie rodzaje autopagnozji i anosognozji występują, gdy proces patologiczny znajduje się po prawej stronie.
Klęska dolnego płatka ciemieniowego może objawiać się zaburzeniem syntezy elementarnych wrażeń lub niemożnością porównania zsyntetyzowanych złożonych wrażeń z „kiedyś w percepcji podobnych
w ten sam sposób, na podstawie wyników, których rozpoznanie następuje ”(V.M. Bekhterev). Przejawia się to naruszeniem dwuwymiarowego zmysłu przestrzennego (grafoestezja) i trójwymiarowego zmysłu przestrzennego (stereognoza) - astereognoza.
W przypadku uszkodzenia stref przedruchowych płata czołowego (pola 6, 8, 44) zwykle występuje ataksja czołowa, w której zaburzona jest synteza impulsów aferentnych (aferentacja kinestetyczna), sygnalizująca położenie części ciała w przestrzeni, które zmiany podczas wykonywanych ruchów.
W przypadku naruszenia funkcji kory przednich części płata czołowego, która ma połączenia z przeciwną półkulą móżdżku (połączenia czołowo-mostowo-móżdżkowe), po przeciwnej stronie ogniska patologicznego występują zaburzenia statokinetyczne (ataksja czołowa). Szczególnie wyraźne są naruszenia późno rozwijających się form statokinetyki - stania w pozycji wyprostowanej i chodzenia w pozycji wyprostowanej. W rezultacie pacjent ma niepewność, chwiejność chodu. Podczas chodzenia jego ciało odchyla się do tyłu. (Znak Hennera) stawia nogi w linii prostej (chód lisa) czasami podczas chodzenia dochodzi do „splatania” nóg. U niektórych pacjentów z uszkodzeniem przednich części płatów czołowych rozwija się osobliwe zjawisko: przy braku porażenia i niedowładu oraz zachowanej zdolności do pełnego poruszania nogami pacjenci nie mogą stać (astazja) i idź (abazja).
Klęska asocjacyjnych stref kory często charakteryzuje się rozwojem klinicznych objawów naruszenia wyższych funkcji umysłowych (patrz rozdział 15).








